 |
| Monodelphis scalops, um representante dos GRUPOS do LESTE |
Grupo Monodelphis dimidiata
Caracteres cranianos nos adultos
Maior comprimento do crânio 38 mm; contorno do crânio em vista dorsal mais estreito que nos grupos anteriores (figs.43 e 44); crista sagital evidente; perfil nasal variando entre quase reto e convexo; forâmen infraorbital situado acima da raiz posterior de P3 ou acima de M1; largura orbital entre um sexto e um oitavo do comprimento do crânio, jugal muito pouco dobrado na região pós-orbital, ângulo de dobramento variando entre 165° e 168°; processo pós-orbital do frontal evidente afastado da constrição intertemporal; crista transversal do alisfenóide evidente; distância (drc) entre o foramen rotundum e canal transverso aproximadamente duas vezes maior que a distância (dtc) entre o canal transverso e o forâmem da carótida; distância (dcc) entre os forâmens das carótidas duas vezes maior que a distância existente entre cada um deles e o canal transverso adjacente; porção orbital do palatino (epp) mais estreita e reduzida, que em outros grupos de espécies; frontal muito próximo do forâmen orbitosfenóide; processo alisfenóide da bula auditiva terminando entre a parte mais anterior da bula e o processo timpânico do periótico (pt); processo timpânico do periótico (pt) em ponta ou levemente arredondado; perfil da borda inferior do ramo horizontal da mandíbula, quase reto na porção central; ângulo interno formado pelo processo coronóide e o ramo horizontal entre 100° e 106°; processo coronóide estreito na ponta; inclinação interna do processo angular entre 100° e 115°, ramo ascendente da mandíbula com inclinação entre 30° e 40° ( Fig. 42).

pré-molares
superiores
diástema entre P1 e P2 e entre P2 e P3; P2 comprimento mesio-distal aproximadamente duas vezes maior que P1 e igual a P3; P2 com largura buco-lingual aproximadamente um terço maior que P1 e pouco menor que P3; P2 com coroa pouco mais baixa que P3 comprimento mesio-distal pouco mais longo que a altura da coroa, P3 com o comprimento mesio-distal aproximadamente igual ao coroa, raízes muito mais expostas em P3 deixando a coroa num plano mais ventral com relação a P1 e P2; P1 com altura da coroa igual a dois terços da mesma em P2 (fig. 45).
diástema entre p1 e p2 e entre p2 e p3; p2 com o comprimento mesio-distal aproximadamente duas vezes ou um terço maior que p1 e um terço maior que p3; p2 com altura da coroa aproximadamente o dobro da mesma em p1 e pouco menor que em p3; raízes mais expostas em p3 como no maxilar (fig. 45).

Molares
superiores
comprimento mesio-distal de M1 pouco maior que de 3; M3 com ectoestilo l mais reduzido que em M1 e M2 ; bacia metaestilar proporcionalmente menor em M3 que em M1 e M2; M4 com ectoestilo j muito reduzido, porção do cíngulo bucal adjacente reduzida (fig. 46).
inferiores
inferiores
comprimento mesio-distal em m4 pouco menor que em m3 (fig. 46)


História taxonômica
De acordo com sua descrição: "Toda a pelagem curta e suave como a do camundongo. Debaixo do olho e muito pouco sobre ele, todo o lado da cabeça e do animal até a cauda, são de cor de canela viva; e algo mais clara na parte inferior do corpo. Sobre o focinho é pardo; e todo o resto, pardo-chumbo, ou melhor uma mistura, porque alguns pêlos tem as pontas esbranquiçadas, que se percebe entre os outros." Segundo um relato de seu amigo Nosedá: Fora encontrada uma fêmea com catorze filhotes e com catorze tetas no ventre. Diferentemente do macho não possuía um odor forte. Comprimento: 6 4/5" (17,3 cm); cauda 2" (5,08 cm). A cor de canela viva, vista nos machos, só se via nos lados da cabeça e no pescoço, nos lados do corpo tornava-se parda por estar fundida com o cinzento de cima. As partes inferiores eram ruivas acaneladas ou acaneladas esbranquiçadas", a qual Azara identifica como seu "colicorto".
Os crânios de Monodelphis brevicaudis e M. dimidiata apresentam diferenças facilmente identificáveis, M. dimidiata possui a distância interorbital mais estreita que M. brevicaudis, isso também vale para M. sorex (=M. dimidiata) que foi baseada em adultos jovens de M. dimidiata, 1 macho e 1 fêmea (em álcool).

Descrição da pelagem
Variação: dos exemplares examinados, MN 1312, macho adulto, e o MN 24605, macho jovem. O macho adulto tem a pelagem mais amarelada que o jovem .
Didelphys unistriata Wagner (1842: 360). Schinz (1844: 505). Waterhouse (1846: 513).
Wagner (1847: 148). Wagner (1855: 249). Giebel (1859: 717).
Microdelphys unistriata: Burmeister (1856: 87).
Didelphys (Peramys) unistriata: Thomas (1888b: 365).
Peramys unistriatus: Cabrera (1919: 43). Miranda-Ribeiro (1936: 417).
Monodelphis unistriata: Cabrera & Yepes (1940: 31). Emmons (1990: 30).
Monodelphis unistriatus: Vieira (1949: 358). Vieira (1955: 349).
Monodelphis (Monodelphis) unistriata: Cabrera (1958: 10).
Localidade tipo: Itararé, São Paulo.
História taxonômica

Caracteres cranianos dos adultos

Variação da forma do crânio entre as espécies:


Dentição
Pré-molares

Molares

Peramys emiliae Thomas (1912: 89)
[Didelphis (]Monodelphis[)] emiliae: Matschie, (1916: 271).
Monodelphis emiliae: Thomas (1920: 283).
P[eramys]. tricolor emiliae: A. Miranda Ribeiro (1936: 407).
P[eramys]. tricolor emilae: C. O. C. Vieira, (1950: 360).
Monodelphis tricolor emiliae: C. O. C. Viera (1955: 350).
Monodelphis (Monodelphis) touan emiliae: Cabrera (1958: 9) .
Monodelphis emiliae: Peterson & Pine (1982: 467).
Monodelphis emiliae: Pine & Handley, Jr. (1984: 239).
Localidade tipo: Boim, rio Tapajós, Pará, Brasil
História taxonômica
Descrição da pelagem

Descrição da pelagem
Comentários
Espécime examinado
Total: 1 s / loc. sexo? (MG 11839).





Mus araneus: Marcgrave (1648: 229).
La musaraigne du Brèsil: Buffon (1767: 160).
Brasilian Shrew: Pennant (1771: 309).
Sorex americanus Muller (1776: 36).
Sorex brasiliensis Erxleben (1777: 127). Schreber (1778: 577). Boddaert (1785: 125). Gmelin (1788: 115). Kerr (1792: 206).
Sorex brasiliensis: Zimmermann (1780:386). Shaw (1800: 535).
Didelphys tristriata Illiger (1815: 107). Kuhl (1820: 63). Fischer (1829: 269). Waterhouse (1841: 107). Schinz (1844: 511). Waterhouse (1846: 518). Wagner (1848: 52).
Didelphis tristriata Schinz (1821: 252).
Didelphys trilineata Lund (1841: 237).
Peramys triistriata: Lesson (1842: 187).
Microdelphys tristriata: Burmeister (1856: 84).
Didelphys (Peramys) americana: Thomas (1888b: 363).
Peramys americanus: Cabrera (1919: 42). Miranda-Ribeiro (1936: 414).
Peramys iheringii: Miranda-Ribeiro (1936: 415) parte.
Monodelphis americana: Cabrera & Yepes (1940: 30). Emmons (1990: 30) parte.
Monodelphis americanus: Vieira (1949: 358 parte. Vieira (1955: 349) parte. Hershkovitz (1959: 338).
Monodelphis (Monodelphis) americana americana: Cabrera (1958: 6) parte.
Monodelphis rubida: Emmons (1990: 30) parte.
Peramys tristriata: Goeldi & Hagmann (1904: 100)
Minuania umbristriata: Miranda-Ribeiro (1936: 422).
M[inuania]. goyana: Miranda-Ribeiro (1936: 419).
Minuania umbristriata: Cabrera & Yepes (1940: 33).
Monodelphis umbristriatus: Vieira (1955: 350).
Monodelphis (Monodelphis) americana americana: Cabrera, (1958: 6) parte.
Monodelphis rubida: Emmons (1990: 30) parte.
História taxonômica

Descrição da pelagem do fenótipo meridional
Descrição da pelagem do fenótipo do Pará
Parte superior da cabeça, lados do focinho, bochechas, ao redor das orelhas, marrom levemente alaranjado, mais alaranjado ao redor das orelhas e nas bochechas, mais escuro no topo da cabeça; parte superior do focinho, enegrecido, as vezes com pequeno segmento preto longitudinal; pescoço, dois terços anteriores do dorso e laterais, marrom levemente amarelado, pelos aristiformes, dorsais e laterais com banda basal cinza e terminal sépia, pelos setiformes dorsais com banda basal cinza e terminal sépia nas faixas e com banda basal cinza, mediana sépia, subterminal amarelo claro e ponta sépia, no resto, pelos setiformes, laterais com banda basal cinza, mediana sépia, subterminal amarelo claro e ponta sépia; o terço posterior do dorso, traseiro e partes superiores dos membros, marrom levemente avermelhado; três faixas pretas ao longo do dorso; faixa central do topo da cabeça a base da cauda, mais difusa na região marrom avermelhada do traseiro; faixas laterais mais curtos, evidentes apenas no terço central do dorso, diluindo-se anteriormente até os ombros e quase imperceptíveis no terço posterior do dorso; queixo, bordas inferiores da boca e garganta amarelo claro rosado, pelos sem banda basal cinza; pescoço inferiormente, peito, barriga e partes internas dos membros, ocre acinzentado, pelos aristiformes, com banda basal cinza, subterminal sépia e ponta amarelo claro, pelos setiformes, com banda basal cinza e terminal amarelo claro; parte superior das patas, marrom claro mais claro nos pés; dedos com escamas; cauda com escamas visíveis e pelos na base, exceto na base inferiormente, marrom escuro superiormente e mais claro inferiormente (fig. 68).
Espécimes examinados
Total: 58. BRASIL - Pará: Mosqueiro, 1 M (MG 3543); Belém, 5 M (MZUSP 11698, MG 318, 327, 1975, 2573), 2 F (MZUSP 9930, MG 2574), 2 ? (MG s/n, MZUSP 8237); 1 M (MN 1304), 2 F (MN 1303, 1305). - Goiás: Veadeiros, 1 M (MN 1313), 1 ? (MN 1314). - Brasília: 1 F (UFPB 2833). - s/loc. 2 M (MZUSP 9415, 10206), 2 F (MN 315, 316).- Paraíba: João Pessoa, 1 M (APO 406), 1 F (UFPB 413); -Pernambuco: Caruarú, 1 M (MN 24544). - Bahia: Ilhéus, 6 M (MN 11075, 11179, 11180, 11485, 11492, 20976); Ilhéus ?, 1 M (MN 11524), 2 F (MN 11483, 11498). - Minas Gerais: Além Paraíba, 2 M (MN 7569, 7571), 1 F (MN 7568); Mariana, 1 M (MZUSP 2193, 1 F (MZUSP 3418); Passos, 2 M (MN 20971, 20972), 3 F (MN 11728, 20973, 20974); Porto Novo, 1 F (MN 7312). - Espírito Santo: Santana, 1 M (MN 1307); - Rio de Janeiro: Itatiaia, 1 F (MZUSP 430); Teresópolis, 1 F (MN 7250). - São Paulo: Paraibuna, 1F (MN 10988); Perus, 1F (MZUSP 11); Piquete, 2 ? (MZUSP 118, 1520); Piracicaba, 4 M (MZUSP 1516, 1518, 1519(a), 1519(b)), 1 F (MZUSP 1523); São Paulo, 2 M (MZUSP 1185, CNFG 1), 1 F (CNFG 2).
A descrição de Thomas corresponde perfeitamente aos exemplares MN 24550, 24551, do Museu Nacional coletados em Jaguaquara, Bahia. Portanto, a única localidade precisa para esta espécie (aqui considerada como uma forma geográfica).
Descrição da pelagem do fenótipo com eritrismo conhecido como "rubida"
Comprimento dos pelos: aristiformes, dorsais 10 mm, laterais 11 mm, ventrais 11 mm: setiformes, dorsais 6 mm, laterais 7 mm e ventrais 4 mm.
Medidas corporais: cabeça e tronco: 126-160 mm; cauda: 65-70 mm; pé: 18-21 mm; orelha: 13-18 mm.




Localidade tipo:"Rio Grande do Sul;" identificada como Brasil, Rio Grande do Sul, Taquara, by Thomas (1888).
História taxonômica

Medidas corporais: cabeça e tronco: 77-110 mm; cauda: 43-50 mm; pé: 14-15 mm; orelha: 12-14 mm.

Espécimes examinados
Total: 21. BRASIL - Rio de Janeiro: Ilha Grande, 2 M (MZUSP 1979, MN 24400); Jacarepaguá, 1 M (MN 24545); Parati, 2 M (MN 8203, 6221), 2 ? (MN s/n, s/n); Restinga da Marambaia, 1 M (MN 1308); Rio de Janeiro, 1 M (MN 10305), 1 F (MN 24546), 1 ? ( MN s/n); Teresópolis, 1 F (MZUSP 1855). - São Paulo: Iguape ?. - Santa Catarina: Colônia Hansa, 3 M (MZUSP 847, 873, 874); Hammonia, 1 M (MZUSP 3417); Joinville, 2 M (MZUSP 3421(a), 3421(b); 1 ? (MZUSP MB 2).
História taxonômica
Descrição da pelagem
Comprimento dos pelos: aristiformes, dorsais 7 mm, laterais e ventrais 8 mm, setiformes, dorsais e laterais 6 mm, ventrais 4 mm.
Medidas corporais: cabeça e tronco: 115-165 mm; cauda: 50-71 mm; pé: 16-25 mm; orelha: 11-16 mm.
Comprimento dos pelos: aristiformes, dorsais 6 mm, laterais 8 mm e ventrais 7 mm, setiformes, dorsais e laterais 5mm e ventrais 4 mm.
Crânios comparação dos adultos velhos e adultos jovens.
Total: 18. BRASIL - Espírito Santo: Alfredo Chaves, 1 F (MZUSP 6674); Venda Nova, 1 M (UFPB 263), 2 F (UFPB 262, 264); - Rio de Janeiro: Parati, 1 M (MN 6419), 1 F (MN 6102); Petrópolis, 1 F (MN 10209); Rio de Janeiro, 1 F (MN 24546); Teresópolis, 5 M (MN 1301, 1302, 7245, 7246, 7247), 4 F (MN 1310, 7233, 7248, 7249); - São Paulo: São Sebastião, 1 F (MZUSP 1528).
LINKS
Monodelphis scalops
:https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEgaYPYIxybesu9cvAOJ0c6hUQYDDyTXmqL38GqykzCdYwgrGOOZhE50unByYVYGWBqJdIyB9ygfjY9DaJGiF3Ewj2ECEvyim0O0Uq00QTwVxbwvM_xhWHho3W5SyGhGEjvQ7cDwJRm8AqvQ/s1600/MonoIhe2.jpg

Comparações cranianas entre as espécies M. dimidiata, M. brevicaudis, M. unistriata:
 |
| Algumas das características cranianas distintivas das espécies do grupo M. dimidiata. Monodelphis dimidiata (=M. sorex). |
 |
| Os crânios do MZUSP 1435 e 1527 de São Lourenço do Sul (31º22’S 51º59’W), RS. Monodelphis dimidiata (=M. sorex). |
 |
| Um outro marsupial que habita a região dos Pampas (gênero Lutreolina), apresenta um crânio com focinho curto e a região interorbital estreita, da mesma forma que em Monodelphis dimidiata, (com os olhos voltados para cima). |
 |
| Monodelphis dimidiata, crânios de um macho e uma fêmea mostrando o pronunciado dimorfismo sexual, ainda que esse exemplar fêmea não seja um adulto velho. Foto restaurada, retirada de: PINE, R. H., DALBY, P. L. & MATSON, J. O., 1985, Ecology, postnatal development, morphometrics, and taxonomic status of the short tailed opossum, Monodelphis dimidiata, an apparently semelparous annual marsupial. Ann. Carnegie Mus. |
Esses dois exemplares representados abaixo foram escolhidos especialmente para mostrar a gama existente nesse grupo de espécies. A figura de cima mostra um crânio com perfil mais reto, silhueta mais alongada (M. brevicaudis) como nas outras espécies do gênero. A figura de baixo mostra um crânio com perfil muito curvo, focinho curto e o ramo mandibular bem dobrado (M. dimidiata), sem dúvida alguma características derivadas. Nenhuma outra espécie de Monodelphis possui tal caráter, aproxima-se mais de M. dimidiata. Temos aqui um extremo importantíssimo sob o ponto de vista evolutivo. As duas espécies desse gênero são perfeitamente distintas entre si. M. brevicaudis apresenta o focinho pouco mais longo, perfil nasal mais reto, processos pós-orbitais dos frontais menores, ramo ascendente da mandíbula menos dobrado e processo coronóide mais alto que M. dimidiata, (fêmea ?) (fig 43 e 44).
 |
| Monodelphis brevicaudis |
 |
| Monodelphis dimidiata (=sorex) |

Monodelphis brevicaudis Olfers. 1818
Micouré cinquième, ou micouré queue courte: Azara (1801: 295)
Colicorto: Azara (1802: 258).
Didelphys brevicaudis: Olfers (1818: 205).
Didelphis tricolor: Desmarest (1820: 260) parte.
Didelphis tricolor: Desmarest (1820: 260) parte.
Didelphis tricolor: Temminck (1827: 52) parte.
D[idelphis] brachyura: Gray (1827: 190) parte.
Didelphis tricolor: Lesson (1827: 212) parte.
Didelphis brachyura: Lund (1837:134; 1842: 135).
D[idelphis]. tricolor: Lund (1839: 237; 1840: 63).
Didelphys brachyura: Burmeister (1854: 141) parte.
D[idelphis]. tricolor: Lund (1839: 237; 1840: 63).
Didelphys brachyura: Burmeister (1854: 141) parte.
D[idelphys]. brachyura: Wagner (1855: 252) parte.
Didelphys (Peramys) henseli: Thomas (1888a: 159). Thomas (1888b 360).
Peramys brevicaudatus: Bertoni (1914: 69).
Peramys henseli: Bertoni (1914: 69).
[Monodelphis] lundi: Matschie (1916: 271).
[Monodelphis] wagneri: Matschie (1916: 272).
Peramys henseli: A. Miranda-Ribeiro (1936: 409).
P[eramys]. d[imidiata]. itatiayae : A. Miranda-Ribeiro (1936: 421)
Monodelphis tricolor paulensis: Vieira (1950: 359). Vieira (1955: 350).
Monodelphis (Monodelphis) henseli: Cabrera (1958: 9).
Monodelphis (Monodelphis) touan paulensis: Cabrera (1958: 9)
Peramys brevicaudatus: Bertoni (1914: 69).
Peramys henseli: Bertoni (1914: 69).
[Monodelphis] lundi: Matschie (1916: 271).
[Monodelphis] wagneri: Matschie (1916: 272).
Peramys henseli: A. Miranda-Ribeiro (1936: 409).
P[eramys]. d[imidiata]. itatiayae : A. Miranda-Ribeiro (1936: 421)
Monodelphis touan: Cabrera & Yepes (1940: 31) parte.
Monodelphis henseli: Cabrera (1940: 32). Vieira (1955: 349) parte.
Monodelphis (Monodelphis) henseli: Cabrera (1958: 9).
Monodelphis (Monodelphis) touan paulensis: Cabrera (1958: 9)
Monodelphis brevicaudis: Hershkovitz (1959: 344).
Monodelphis henseli: Ruschi (1965: 3).
Mondelphis dimidiata itatiayae: Pine (1979: 495).
Monodelphis sorex: Voss (1975: 9).Emmons (1990: 29).
Localidade tipo: perto de San Ignacio Guazú, Misiones, Paraguay
Mondelphis dimidiata itatiayae: Pine (1979: 495).
Monodelphis henseli: Pine (1979: 495).
[Monodelphis] sorex: Pine & Handley, Jr. (1984: 242)parte .
Monodelphis brevicaudata: Emmons (1990: 27) parte.
Localidade tipo: perto de San Ignacio Guazú, Misiones, Paraguay
História taxonômica
Félix de Azara (1801: 295) foi o primeiro autor a descrever esta espécie no seu "Micoure cinquième, ou micouré à queue courte". Redescrito em espanhol em Azara (1802: 258). Em sua versão espanhola cita que teve dois machos coletados no campo sob os paralelos 27º 34' (o extremo sul do Paraguai, Itapuá). O comprimento do corpo: 6 ¾ (17,2 cm); cauda 2 ¼ (5,7 cm); orelha: 4" (0,85 cm). Um amigo de Azara chamado Nosedá teve um exemplar macho engaiolado, com comprimento de 8 ½ polegadas (21,7 cm), e com as mesmas proporções do anterior.
 |
| Félix de Azara foi militar, engenheiro, explorador, cartógrafo, antropólogo, humanista e naturalista. |
De acordo com Hershkovitz (1959: 344), Olfers (1818: 205) propôs o nome Didelphys brevicaudis (até então nomem nudum) com base no "Micouré à queue courte" de Azara, sendo assim o primeiro nome válido da espécie.
Vários outros autores referem-se posteriormente ao animal descrito por Azara. Desmarest (1820: 260) e Temminck (1827: 52) em Didelphis tricolor (Geoffroy). Gray (1827: 190) em D. brachyura (Pallas), "Inhabits Cayena and Paraguai". Lesson (1827: 212) coloca-o em Didelphis tricolor (Geoffroy), "Micouré cinquieme d'Azara". Lund (1839: 237; 1840: 63; 1841: 236 e 1842: 135) citando D. tricolor (Geoffroy) ou D. brachyura (Pallas) refere-se ao Colicorto de Azara. Wagner (1843: 50) no D. tricolor (Geoffroy) cita o animal de Azara e a localidade "Paraguay". Burmeister (1854: 142) em Didelphys brachyura (Schreber ), relaciona-o ao Colicorto.
Vários outros autores referem-se posteriormente ao animal descrito por Azara. Desmarest (1820: 260) e Temminck (1827: 52) em Didelphis tricolor (Geoffroy). Gray (1827: 190) em D. brachyura (Pallas), "Inhabits Cayena and Paraguai". Lesson (1827: 212) coloca-o em Didelphis tricolor (Geoffroy), "Micouré cinquieme d'Azara". Lund (1839: 237; 1840: 63; 1841: 236 e 1842: 135) citando D. tricolor (Geoffroy) ou D. brachyura (Pallas) refere-se ao Colicorto de Azara. Wagner (1843: 50) no D. tricolor (Geoffroy) cita o animal de Azara e a localidade "Paraguay". Burmeister (1854: 142) em Didelphys brachyura (Schreber ), relaciona-o ao Colicorto.
Thomas (1888a: 159) descreveu Didelphys (Peramys) henseli baseado em cinco exemplares coletados em Taquara, Rio Grande do Sul: "Tamanho intermediário entre o de D. brevicaudata e os grupos D. sorex e Iheringi. Cor cinza escuro grisalho ao longo de toda a superfície superior, ruivo profundo nas laterais e na barriga. Orelhas pequenas, atingindo quando caídas apenas a metade da distância na direção do olho. Mamas cerca de vinte e cinco em número, cinco centrais e cerca de dez pares laterais". Pouco tempo depois, Thomas (1888b: 360) redescreveu essa espécie, e as características por ele apresentadas não deixam dúvidas quanto a sua correspondência com o Colicorto de Azara (M. brevicaudis).
Bertoni (1914: 69) lista duas espécies de Peramys no Paraguai P. brevicaudatus e P. henseli. Provavelmente observando diferenças relacionadas a idade dos exemplares, identificou duas espécies em seu material.
Paul Matschie (1916: 271-272), numa listagem, propõe dois nomes novos baseado em citações de outros autores, [Monodelphis] lundi para D[idelphis]. tricolor: Lund (1841: 236, 1842: 135) de Lagoa Santa e [Monodelphis] wagneri, com base na referência de Wagner (1855: 252) ao "Micouré à queue court" de Azara, do Paraguai, ambos sinônimos juniores de M. brevicaudis de Olfers.
Miranda-Ribeiro (1936: 421) descreve P[eramys]. d[imidiata]. itatiayae baseado em um exemplar macho de Campo Belo, Rio de Janeiro (Itatiaia). Pine (1979: 495) examinou o holótipo desta subespécie, que encontra-se no National Museum of Natural History, e não encontrou justificativas para o reconhecimento da suposta subespécie nova, identificando-a como M. henseli. Provavelmente, diferenças relativas a idade do exemplar e o desconhecimento de tais variações levaram Miranda-Ribeiro a propor a sua subespécie.
Cabrera & Yepes (1940: 31) em Monodelphis touan, inclui na distribuição dessa espécie, "Paraguay" e a seguir cita Bertoni. Portanto, ele considerou Monodelphis brevicaudis Olfers, como sinônimo da espécie da Guiana Francesa e imediações. Nesse mesmo trabalho considerou M. henseli uma espécie válida. As antigas confusões, originadas pelas afirmações de Azara sobre o seu colicorto, comparando-o ao Touan de Buffon, repercutiram por longo tempo.
Vieira (1950: 359) propõe Monodelphis tricolor paulensis baseado em um exemplar macho adulto de Pirituba, São Paulo. As mesmas semelhanças, já observadas por outros autores, existentes entre M. brachyura Pallas e M. brevicaudis Olfers, levou esse autor a considerá-las apenas como subespécies diferentes. Vieira considerou essa subespécie como próxima de "P[eramys]. tricolor emiliae", diferindo desta por possuir "todo alto da cabeça até a ponta do focinho a mesma coloração do dorso, cinza-esbranquiçado e a região da base da cauda, ferrugíneo muito apagado".
Pine & Handley (1984:242) incluem [Monodelphis] henseli como sinônimo junior de [Monodelphis] sorex, numa lista provisória. Emmons (1990: 29) provavelmente seguindo a postura desses autores tomou a mesma atitude. Considero aqui M. sorex, como sinônimo de M. dimidiata.
Emmons considera também "N[orthern]. Argentina" como parte da distribuição de M. brevicaudata. Nessa referência devem estar embutidas às antigas confusões existentes com relação à presença de "D. brachyura" Schreber no Paraguai, que se referem na verdade à Monodelphis brevicaudis Olfers, 1818.
Bertoni (1914: 69) lista duas espécies de Peramys no Paraguai P. brevicaudatus e P. henseli. Provavelmente observando diferenças relacionadas a idade dos exemplares, identificou duas espécies em seu material.
Paul Matschie (1916: 271-272), numa listagem, propõe dois nomes novos baseado em citações de outros autores, [Monodelphis] lundi para D[idelphis]. tricolor: Lund (1841: 236, 1842: 135) de Lagoa Santa e [Monodelphis] wagneri, com base na referência de Wagner (1855: 252) ao "Micouré à queue court" de Azara, do Paraguai, ambos sinônimos juniores de M. brevicaudis de Olfers.
Miranda-Ribeiro (1936: 421) descreve P[eramys]. d[imidiata]. itatiayae baseado em um exemplar macho de Campo Belo, Rio de Janeiro (Itatiaia). Pine (1979: 495) examinou o holótipo desta subespécie, que encontra-se no National Museum of Natural History, e não encontrou justificativas para o reconhecimento da suposta subespécie nova, identificando-a como M. henseli. Provavelmente, diferenças relativas a idade do exemplar e o desconhecimento de tais variações levaram Miranda-Ribeiro a propor a sua subespécie.
Cabrera & Yepes (1940: 31) em Monodelphis touan, inclui na distribuição dessa espécie, "Paraguay" e a seguir cita Bertoni. Portanto, ele considerou Monodelphis brevicaudis Olfers, como sinônimo da espécie da Guiana Francesa e imediações. Nesse mesmo trabalho considerou M. henseli uma espécie válida. As antigas confusões, originadas pelas afirmações de Azara sobre o seu colicorto, comparando-o ao Touan de Buffon, repercutiram por longo tempo.
Vieira (1950: 359) propõe Monodelphis tricolor paulensis baseado em um exemplar macho adulto de Pirituba, São Paulo. As mesmas semelhanças, já observadas por outros autores, existentes entre M. brachyura Pallas e M. brevicaudis Olfers, levou esse autor a considerá-las apenas como subespécies diferentes. Vieira considerou essa subespécie como próxima de "P[eramys]. tricolor emiliae", diferindo desta por possuir "todo alto da cabeça até a ponta do focinho a mesma coloração do dorso, cinza-esbranquiçado e a região da base da cauda, ferrugíneo muito apagado".
Pine & Handley (1984:242) incluem [Monodelphis] henseli como sinônimo junior de [Monodelphis] sorex, numa lista provisória. Emmons (1990: 29) provavelmente seguindo a postura desses autores tomou a mesma atitude. Considero aqui M. sorex, como sinônimo de M. dimidiata.
Emmons considera também "N[orthern]. Argentina" como parte da distribuição de M. brevicaudata. Nessa referência devem estar embutidas às antigas confusões existentes com relação à presença de "D. brachyura" Schreber no Paraguai, que se referem na verdade à Monodelphis brevicaudis Olfers, 1818.
Os crânios de Monodelphis brevicaudis e M. dimidiata apresentam diferenças facilmente identificáveis, M. dimidiata possui a distância interorbital mais estreita que M. brevicaudis, isso também vale para M. sorex (=M. dimidiata) que foi baseada em adultos jovens de M. dimidiata, 1 macho e 1 fêmea (em álcool).
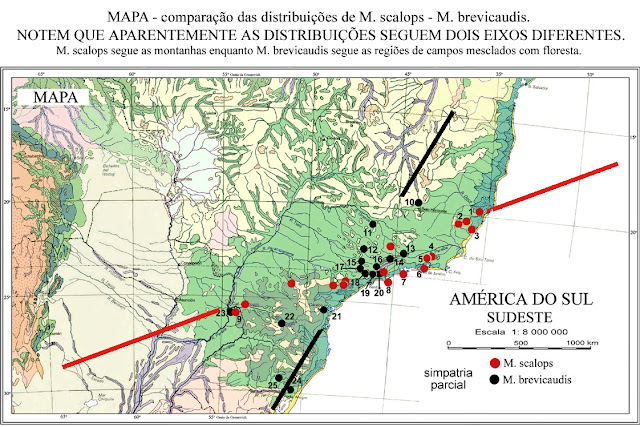
Recentemente foi publicado um trabalho envolvendo análise molecular, considerando as duas espécies desse grupo como formas geográficas de uma única espécie: M. dimidiata. VILELA, J. F. - RUSSO,C.A. De M. & OLIVEIRA, J. A. De ( 2010 ). Esse trabalho falhou por considerar uma simpatria desses "fenótipos" (que não existe), os exemplares utilizados como prova de simpatria das formas, são apenas variações locais de M. brevicaudis (possuem uma larga banda cinza na base dos pelos ventrais, isso não existe em M. dimidiata).Vejam a comparação das formas na figura abaixo. Por outro lado, existe uma distribuição geográfica desses "fenótipos" e terão que ser nomeados por questões de referência. M. dimidiata é a forma dos Pampas e M. brevicaudis está associado as áreas da Floresta de Araucaria. M. sorex é sinônimo junior de M. dimidiata, vinha sendo considerada como sinônimo de M. brevicaudis, mas isso é um erro. Existem evidências de simpatria em Taquara no Rio Grande do Sul e na borda da Serra Gaúcha.


 |
| Monodelphis brevicaudis, Fazenda Marimbondo (22°21’47”S, 44°35’47,47”W), mun. de Itatiaia, Rio de Janeiro - BRASIL Foto: Pessôa-Modesto-Bergallo |
 |
| Monodelphis brevicaudis, Mogi das Cruzes, São Paulo, Brasil. foto de Antonio Wuo. |
 |
| Monodelphis brevicaudis, Mogi das Cruzes, São Paulo, Brasil. foto de Antonio Wuo. |
 |
| Monodelphis brevicaudis, Mogi das Cruzes, São Paulo, Brasil. foto de Antonio Wuo. |

Coloração geral: cinza e marrom-alaranjado. Parte superior do focinho, entre os olhos, topo da cabeça e dorso cinza-grisalhos; lados do focinho, ao redor dos olhos, bochechas, ao redor da orelhas, lados do pescoço, laterais do corpo, traseiro, partes superiores dos membros e base da cauda marrons-alaranjadas, mais alaranjado nas bochechas, ao redor das orelhas e mais avermelhado no traseiro.
 |
| Monodelphis brevicaudis, Pantanal - BRASIL |
 |
Monodelphis brevicaudis, Campo Belo do Sul, SC - BRASIL foto: (c) Felipe Peters |
Descrição da pelagem
Coloração geral cinza e marrom-alaranjado. Parte superior do focinho, entre os olhos, topo da cabeça e dorso cinza-grisalhos, pelos aristiformes com banda basal cinza e terminal preta, setiformes com banda basal cinza, mediana preta subterminal branca-amarelada e ponta preta; lados do focinho, ao redor dos olhos, bochechas, ao redor da orelhas, lados do pescoço, laterais do corpo, traseiro, partes superiores dos membros e base da cauda marrons-alaranjadas, mais alaranjado nas bochechas, ao redor das orelhas e mais avermelhado no traseiro, pelos aristiformes com banda basal cinza subterminal preta e ponta alaranjada setiformes com banda cinza, subterminal laranja-clara e ponta preta; limite entre o cinza dorsal e o marrom-alaranjado das laterais, mal definido; queixo, bordas inferiores da boca e garganta, alaranjados, banda basal cinza; parte inferior do pescoço. peito, barriga e partes internas dos membros cinzas-alaranjados, pelos com banda basal, cinza e banda terminal, branca-alaranjada; cauda com escamas cobertas por pelos pequenos dorsalmente, marrom-escuro, ventralmente, mais cinza nas partes superiores das patas, alaranjadas-claras (fig. 47).
Variação: o cinza dorsal pode ser levemente ou fortemente alaranjado, o marrom-alaranjado das partes laterais pode também ser fortemente alaranjado, principalmente ao redor das orelhas. As partes inferiores do corpo podem ser mais acinzentadas, ou alaranjadas, dependendo da relação entre as larguras dos comprimentos das bandas basais e terminais dos pelos. Jovens, mais avermelhados, o MN 8270, exibe um avermelhado geral por todo corpo, inclusive nos pelos pequenos da cauda. Um exemplar adulto, MN 24549, não possui banda basal cinza nos pelos das partes internas dos membros, barriga e partes inferior do pescoço.



Monodelphis brevicaudis, Bom Jesus, RS (animal jovem)
Foto: (c) Felipe Peters
Foto: (c) Felipe Peters

Comprimento dos pelos: aristiformes, dorsais 9 mm, laterais 11 mm, ventrais 10 mm, setiformes, dorsais 7 mm, laterais 8 mm e ventrais 7 mm.
Medidas corporais: cabeça e tronco: 100-160 mm; cauda: 52-85; pé: 13-17 mm; orelha: 10-13 mm
Espécimes examinados
Total 22. BRASIL - Minas Gerais: Passos, 1 F (MN 20975); - Rio de Janeiro: Itatiaia.- 2 M (MN 24552, 24553); - São Paulo: Amparo, 1 M (MZUSP 6612), 1 F (MZUSP 6611); Jundiaí, 1 ? (MZUSP s/n); Mogi das Cruzes, 1 M (MZUSP 4146), 1 ? (MZUSP s/n); Pinhal, 1 ? (MZUSP 7434); Piquete, 1 F (MN 23869), 2 ? (MZUSP 25, 1526); Pirituba 1 M (MZUSP 2557); São Paulo, 1 M (MZUSP 7671);. Jacareí, 2 M (CFL s/n, s/n); Salesópolis, 1 M (MN 24549); - Paraná: Bituruna, 1 ? (MN s/n); Piraquara, 1 ? (UFP CA2 - 172); PARAGUAI: Puerto Jaguarazapá, 1 M (MZUSP 2823); ARGENTINA: Puerto Bertoni, 1 F (MZUSP 2820); s/loc 1 M (MN 8270).
 |
Essas duas espécies podem ser facilmente diferenciadas pelos traseiros. Monodelphis dimidiata possui um traseiro cinza, enquanto M. brevicaudis é avermelhado ou alaranjado. |
 |
| Foto: Enrique González |
Monodelphis dimidiata Wagner 1847
Didelphis brachyura: Temminck (1827: 53) parte. Waterhouse (1838: 64). Waterhouse (1839: 97). Waterhouse (1841: 111). Wagner (1843: 51) parte. Schinz (1844: 519) parte. Waterhouse (1846: 522).Burmeister (1854: 141).Wagner (1855: 251) parte. Giebel (1859: 716) parte. Burmeister (1879: 194).
D.[idelphys]. dimidiata: Wagner (1847: 151).
Microdelphys sorex: Hensel (1872: 122).
Didelphys (Peramys) dimidiata: Thomas (1888b: 355).
Didelphys (Peramys) sorex: Thomas (1888b: 362).Minuania dimidiata: Cabrera (1919: 43). Cabrera & Yepes (1940: 33).
Monodelphis fosteri: Thomas (1924: 586).
M[inuania] dimidiata : Miranda-Ribeiro (1936: 419).
Monodelphis sorex: Cabrera & Yepes (1940: 32). Vieira (1955: 349)
Minuania fosteri: Cabrera & Yepes (1940: 34).
Monodelphis henseli: Vieira (1955: 349) parte.
Monodelphis dimidiatus: Vieira (1955: 350).
Microdelphys brachyura: Burmeister (1856: 86) parte. Hensel (1872: 122).
Monodelphis (Monodelphis) sorex: Cabrera (1958: 9).
Monodelphis (Minuania) dimidiata: Cabrera (1958: 10) parte.
Monodelphis (Minuania) fosteri: Cabrera (1958:11).
Monodelphis dimidiata: Reig (1964: 207). Fornes & Massoia (1965: 102).
M[onodelphis]. dimidiata: Emmons (1990: 29).
Localidade tipo: Uruguay, Maldonado, "Maldonado am la Plata".
História taxonômica
Esta espécie foi descrita pela primeira vez por Temminck (1827: 53), como Didelphis brachyura de Schreber. Apesar das diferenças existentes entre as duas descrições, por ser D. brachyura a única espécie do gênero que caracterizava-se pelo curto comprimento da cauda, o pouco conhecimento da época não o permitiu ir além. Em sua descrição Temminck dá como lugar de origem Suriname (provavelmente referindo-se às primeiras citações de D. brachyura), um exemplar de Monte-Video coletado pelo capitão Freycinet (o qual deve certamente ter sido usado em sua descrição) e outro do Brasil, enviado por [Auguste] Saint-Hilaire.
A descrição de Temminck concorda com M. dimidiata no aspecto da pelagem, na coloração das patas (esbranquiçadas), na caracterização das orelhas (bem pequenas, arredondadas), focinho curto e curvo, e na proporção corpo-cauda. Porém a descrição que ele fez da pilosidade da cauda é diagnóstica.
 |
The zoology of the voyage of H.M.S. Beagle. By George R. Waterhouse, 1838.
Edited and superintended by Charles Darwin. London: Smith Elder and Co.
|
Infelizmente Didelphis brachyura ficou mais conhecida pela descrição de Temminck, do que pela descrição original de Schreber (de 1778). Pois foi seguido por Waterhouse (1838, 1839, 1841 e 1846), Wagner (1843 e 1855), Schinz (1844: 519), Burmeister (1859 e 1879) e Giebel (1859: 716). Essa substituição chegou a tal ponto, que Wagner (1843:51) no Supplementum III-IV do Die Saugthiere de Schreber, usou a descrição de Temminck ao invés da original.
Apesar disso foi a próprio Wagner (1847: 151), o primeiro a chamar a atenção sobre o desacordo existente entre as descrições que Schreber, Geoffroy, Temminck e Waterhouse fizeram de D. brachyura. Propondo numa nota de roda-pé o nome D. dimidiata para a D. brachyura de Waterhouse, que foi descrita com base num exemplar de Maldonado, Uruguai.
Thomas (1924: 586) descreve Monodelphis fosteri com base em um exemplar juvenil de Caleufú, La Pampa, Argentina, comparando-o com M dimidiata, conclui que a coloração dorsal é muito mais clara e os lados pouco mais amarelados que o dorso, o ventre só levemente amarelado assim como as bochechas. O crânio e os dentes como em M. dimidiata.
Cabrera (1958: 11) trata a espécie de Thomas como Monodelphis (Minuania) fosteri, colocando-a junto com M. dimidiata num mesmo subgênero.
Reig (1964: 207) examinando vinte e nove exemplares da Província de Buenos Aires, Argentina, chega a conclusão que os exemplares jovens de M. dimidiata correspondem em colorido ao exemplar de Thomas, considerando Monodelphis fosteri como sinônimo de M. dimidiata, questionando ainda a distinção do subgênero de Cabrera ("no nos parece suficiente fundamentada"). Em seu trabalho cita um fato bastante interessante relacionado às coletas por ele realizadas. No mês de janeiro foram capturados exemplares adultos de grande tamanho, 3 machos com comprimento total: 209, 221 e 212 mm e 3 fêmeas com: 168, 186 e 173 mm), enquanto nos meses de primavera, outono e inverno foram capturados vinte e três exemplares todos jovens, com comprimento total entre 103-135 mm.
Coloração geral cinza-amarelada e alaranjada-clara. Parte superior do focinho, entre os olhos, topo da cabeça, parte superior do pescoço e dorso até a base da cauda cinza-grisalhos, levemente ou fortemente amarelados, pêlos aristiformes com banda basal marrom-acinzentada ou cinza e terminal sépia ou preta, pêlos
setiformes, com banda basal cinza, mediana sépia ou sem banda mediana, a subterminal branca-amarelada e ponta preta; laterais do focinho, ao redor dos olhos, bochechas, ao redor das orelhas, lados do pescoço, laterais do corpo e partes superiores dos membros alaranjados, mais vivo nas bochechas, ao redor das orelhas e pescoço, pêlos aristiformes com banda basal cinza, subterminal preta ou marrom e ponta amarela ou branca-amarelada, pêlos setiformes com banda basal cinza e terminal amarela; cor do dorso se misturando gradualmente com a cor das laterais; queixo, bordas da boca, garganta e partes internas dos membros amarelos-claros, pêlos sem banda basal cinza; parte inferior do pescoço, peito, barriga cinzas-amarelados ou alaranjados-claros, pêlos com banda basal cinza ou inteiramente alaranjados; cauda com pêlos iguais aos do corpo apenas na base, o resto com pêlos curtos, pêlos da base, dorsalmente cinzas-amarelados, ventralmente alaranjados, pêlos curtos amarelados; o resto da cauda, dorsalmente, cinza-amarelado, ventralmente amarelado; partes superiores das patas amarelas-claras (fig. 47).
Variação: dos exemplares examinados, MN 1312, macho adulto, e o MN 24605, macho jovem. O macho adulto tem a pelagem mais amarelada que o jovem .
 |
| Monodelphis dimidiata - exemplar jovem, coloração diferente dos adultos Foto do grupo: GIECO da Univ. Nac. de Río Cuarto, Córdoba, Argentina. |
 |
| Monodelphis dimidiata - Carrasco, Uruguay - FOTO : Gustavo Casás |
 |
| Monodelphis dimidiata - Carrasco, Uruguay - FOTO : Gustavo Casás |
 |
| Monodelphis dimidiata - Carrasco, Uruguay - FOTO : Gustavo Casás |
 |
| Monodelphis dimidiata - Carrasco, Uruguay - FOTO : GustavoCasás |
 |
| Monodelphis dimidiata - Carrasco, Uruguay - FOTO : Gustavo Casás |
  |
| Monodelphis dimidiata do Uruguai Foto: Enrique González |
 |
| Monodelphis dimidiata - URUGUAI Foto: Enrique González |
 |
| Crânio de um animal adulto, com caninos bem desenvolvidos e crista sagital bem formada, Uruguai |
 |
| Crânios de Monodelphis dimidiata (vista dorsal) à esq. um macho e à dir. uma fêmea. Sem a menor dúvida, a redescrição de M. sorex Hensel, 1872 feita por Thomas, refere-se à um adulto jovem de M. dimidiata |
 |
As diferenças cranianas existentes entre os adultos mais velhos dessas espécies também se evidenciam nos mais jovens. Particularmente a distância (largura) interorbital. Sem dúvida são espécies distintas !
Temos muito o que aprender sobre comparações de sequências ! Muito mesmo ! |
 |
| Monodelphis dimidiata COLICORTO PAMPEANO - San Antonio de Areco, Buenos Aires, ARGENTINA FOTO de Manuco Mercante Site de origem http://www.fotonat.org/altaresol.php?image_id=54251 |
Comprimento dos pêlos: no MN 24605, aristiformes, dorsais e laterais 9 mm, ventrais 10 mm, setiformes, dorsais e laterais 7 mm, ventrais 10 mm. No 1312 , aristiformes, dorsais 7 mm, laterais e ventrais 8 mm, setiformes, dorsais e laterais mm e ventrais 7 mm.
Medidas corporais: cabeça e tronco: 72-151 mm; cauda: 37-80 mm; pé: 11-17 mm; orelha: 10-12 mm.
Espécimes examinados
Total: 6. BRASIL - Rio Grande do Sul: São Lourenço, 1 M (MZUSP 1527), 1 M (MZUSP 1435), 1 ? (MZUSP 1540); s/localidade 1 M (MN 1312), 1 ? (MZUSP 2563). ARGENTINA - Buenos Aires: Pergamino, 1 M (MN 24605).
 |
| Monodelphis dimidiata (=sorex) Hensel 1872 |
APARECEU UMA DÚVIDA quanto ao sexo do exemplar ! Se for uma fêmea a hipótese do dimorfismo é a melhor escolha. Nesse caso M. sorex pode ser um sinônimo junior de M. dimidiata.
Microdelphys sorex: Hensel (1872: 122)
Didelphys (Peramys) sorex: Thomas (1888b: 362).
Monodelphiops sorex: Matschie (1916: 261).
Monodelphis sorex: Cabrera & Yepes (1940: 32). Vieira (1955: 349)
Microdelphys sorex: Hensel (1872: 122)
Didelphys (Peramys) sorex: Thomas (1888b: 362).
Monodelphiops sorex: Matschie (1916: 261).
Monodelphis sorex: Cabrera & Yepes (1940: 32). Vieira (1955: 349)
Hensel (1872: 122) descreveu Microdelphys sorex
baseado em exemplares do Rio Grande do Sul. Todas as características
apontadas correspondem às características dos exemplares jovens de M. dimidiata
como já citei anteriormente, os adultos jovens de vários didelfídeos
diferem tanto dos adultos velhos, que foram muitas vezes classificados
como espécies diferentes. Dentre os exemplares examinados por mim,
observei uma grande variação na forma do crânio (fig.20 E).
Como existe uma variação significativa no formato do crânio durante o
crescimento, fiquei confuso e optei pela variação. Mas depois de um
trabalho feito no Rio Grande do Sul, sobre identificação de pequenos
mamíferos com base na microestrutura dos pelos, ABREU, M. S. Lobato,
CHRISTOFF, A. U. & VIEIRA, E. M., 2011, que resultou na
identificação de 3 diferentes padrões, indicando a existência de uma
possível terceira espécie, estou reavaliando.
"Se o crânio que antes identifiquei como M. dimidiata, for de um macho adulto e velho, pode ser na verdade M. sorex de Hensel. Nesse caso essa espécie seria válida"
 |
| Monodelphis unistriata Wagner (1842) |
Didelphys unistriata Wagner (1842: 360). Schinz (1844: 505). Waterhouse (1846: 513).
Wagner (1847: 148). Wagner (1855: 249). Giebel (1859: 717).
Microdelphys unistriata: Burmeister (1856: 87).
Didelphys (Peramys) unistriata: Thomas (1888b: 365).
Peramys unistriatus: Cabrera (1919: 43). Miranda-Ribeiro (1936: 417).
Monodelphis unistriata: Cabrera & Yepes (1940: 31). Emmons (1990: 30).
Monodelphis unistriatus: Vieira (1949: 358). Vieira (1955: 349).
Monodelphis (Monodelphis) unistriata: Cabrera (1958: 10).
Localidade tipo: Itararé, São Paulo.
 |
Foto do tipo de Monodelphis unistriata (cedida por Ronald Pine)
|
Wagner (1842: 360) descreve Didelphys unistriata baseado num exemplar coletado por Natterer em Itararé, São Paulo. Tamanho: corpo: 5 1/4" (13,4 cm); cauda: 2 1/2" (6,4 cm).
"Didelphys supra ferruginea, cano-mixta, subtus rufescens, stria dorsali obscuriori, cauda abbreviata pilosiuscula".
A espécie foi redescrita em maiores detalhes por Wagner (1847: 148) (entre outras medidas o tamanho da orelha foi fornecido: 10 mm.). É interessante notar que desde a sua descoberta nunca foi encontrada, permanecendo um único exemplar conhecido, até recentemente.
"Didelphys supra ferruginea, cano-mixta, subtus rufescens, stria dorsali obscuriori, cauda abbreviata pilosiuscula".
A espécie foi redescrita em maiores detalhes por Wagner (1847: 148) (entre outras medidas o tamanho da orelha foi fornecido: 10 mm.). É interessante notar que desde a sua descoberta nunca foi encontrada, permanecendo um único exemplar conhecido, até recentemente.
Didelphys unistriata. Wagner. Archive für Naturgesch. 1842. p.360, Abh. d. Münchn. Akadem. V. p. 148.
"O aspecto dessa espécie é muito semelhante ao de D. glirina, mas o tamanho é um pouco menor, e a cauda de uma natureza diferente. O corpo é alongado e com pernas curtas. A cabeça é um pouco grossa e curta, o nariz é sulcado, as vibrissas, curtas e finas; as orelhas pequenas, não se projetando além do topo da cabeça, cortadas na parte inferior, nuas, com um delicado toque de pelos. As patas da frente são mais robustas que as traseiras, que são estreitas. A cauda não tem nem a metade do comprimento do corpo, na base é um pouco coberta com o pêlo das costas, no resto densamente coberta com pelos salientes até a ponta. Difere das caudas de D. glirina e velutina, por não ser tão grossa e nem cônica na base, o pelo da cauda não aderido, é mais áspero, e suas pontas não são como naquelas, esticadas em linha reta, mas são encurvadas como ganchos, na parte inferior. Com base nessas curvaturas, parece quase certo, que a cauda de D. unistriata se destina a agarrar, o que também se confirma pelo fato do pelo na parte inferior da ponta da cauda estar mais aderido. O escroto é densamente peludo. A fêmea ainda não é conhecida, mas provavelmente não possui bolsa. O pelo é muito curto e não parece macio. A cor da parte superior é marrom-ferrugem avermelhado com manchas brancas finas. As partes laterais inteiras e o lado de baixo inteiro são monocromáticos e com uma cor de ferrugem clara avermelhada, que recorta bastante a cor das costas e fica um pouco mais claro na parte de baixo. Da cernelha até a raiz da cauda, corre uma faixa estreita levemente recuada, de uma sólida cor vermelho-ferrugem. Os pêlos manchados da parte superior da cabeça e das costas são grisalhos na parte inferior, depois amarelados com pontas marrom-ferrugem avermelhado; os pelos nas laterais e no abdômen são de cor sólida, mais claros perto das raízes, a cabeça não tem distinção especial; as vibrissas são pretas, as cerdas das bochechas são brancas. Os membros são cor de ferrugem como os lados, mas do lado de fora do traseiro a cor pontilhada das costas diminui um pouco. As garras são branco amarelado com uma mancha escura. A cauda é marrom escuro enferrujado por cima, pelos amarelo-ferrugem, sujos por baixo. O escroto preto é coberto com pelos amarelo-ferrugem. As solas parecem cor de carne em vida".
Corpo......................................5" 7'" Do nariz a orelha....................1" 1"'
Cauda.....................................2" 5'" Orelha...................................0" 5'"
Do nariz ao olho.......................0" 6 1/2'" Patas traseiras.......................0" 6 3/4"'
Encontrado por Natterer em Itararé no Brasil.
Thomas (1888b: 365) e Miranda-Ribeiro (1936: 417) examinaram o holótipo de M. unistriata e fizeram descrições próprias e medidas. Estas concordam de modo geral, com exceção da medida da orelha dada por Thomas (5,5 mm), talvez tenha sido medida pelo topo da cabeça e não pela base do tragus.
Recentemente (2013), Pine, Flores & Bauer redescreveram a espécie, utilizando um segundo exemplar, depois de permanecer um longo tempo conhecida apenas pelo tipo coletado por Natterer. Pine, Flores & Bauer. 2013. The second known specimen of Monodelphis unistriata (Wagner) (Mammalia: Didelphimorphia), with redescription of the species and phylogenetic analysis.
(estou adicionando comentários e vou inserir parte dessa re-descrição)
 |
| Vista lateral do tipo mostrando a coloração. Tanto a pelagem quanto o crânio (infelizmente fragmentado) mostram afinidades com o grupo M. dimidiata |

 |
| Monodelphis obscura Versão clareada da foto de Douglas Machado do Parq. Est. Chandless, estado do ACRE - BRASIL. |
Grupo Monodelphis emiliae
Caracteres cranianos dos adultos
Maior comprimento do crânio 38 mm; forma do crânio (fig. 37); crista sagital presente; perfil nasal levemente convexo; forâmen infraorbital, acima da raiz posterior de P3; largura orbital, aproximadamente um sexto do comprimento do crânio; jugal dobrado na região orbital, com um ângulo interno de aproximadamente 160°; processo pós-orbital do frontal evidente; crista transversal do alisfenóide evidente; distância (drt) entre o foramen rotundum e o canal transverso aproximadamente duas vezes maior que a distância (dtc) entre o canal transverso e o forâmem da carótida; distância (dcc) entre os forâmens das carótidas aproximadamente o triplo da distância existente entre cada um deles e o canal transverso adjacente; porção orbital do palatino (epp) levemente alargada; processo alisfenóide (pa) da bula auditiva terminando próximo ao processo timpânico do periótico, bula quase fechada na região mesio-ventral; processo timpânico do periótico (pt) arredondado; perfil da borda inferior do ramo horizontal da mandíbula levemente curvo; ângulo interno formado pelo processo coronóide e o ramo horizontal, aproximadamente 120°; processo coronóide estreito; inclinação interna do processo angular aproximadamente 110°; ramo ascendente da mandíbula com inclinação de aproximadamente 15° (fig.36).

O grupo Monodelphis emiliae é composto por 2 espécies, cada uma delas ocorrendo em uma das margens do rio Madeira. Monodelphis obscura na margem esquerda e M. emiliae na direita. Aparentemente M. obscura é uma espécie menor que M. emiliae, o crânio parece mais curto. Os poucos exemplares examinados ainda não permitem uma afirmação nesse sentido.
 |
Crânios das duas únicas espécies do GRUPO Monodelphis emiliae comparados em vista lateral.
O crânio de M. obscura é um exemplar do MZUSP, um macho, com número de campo MJ 46,
Estação Ecológica Estadual Antônio Mojica Nava, Rondônia, BR e está em escala aproximada.
|

Crânios das 2 espécies do GRUPO Monodelphis emiliae comparados em vista ventral ( M. obscura - exemplar do MZUSP, o mesmo mostrado acima, em escala aproximada ). M. obscura aparentemente é uma espécie menor do que M. emiliae. Vejam este crânio da direita, não é como um animal jovem de M. emiliae, é mais robusto, possui um palato proporcionalmente mais estreito e bulas auditivas menores.

Dentição
Pré-molares
Superiores - diástema entre P1 e P2; P1 com o comprimento mesio-distal igual a dois terços de P2; P3 com comprimento mesio-distal pouco maior que P2; maior largura de P3 pouco maior que a maior largura de P2 (fig. 39).
Inferiores - diástema entre pl e p2 e entre p1 e p3, o diástema anterior maior que o posterior; comprimento mesio-distal de p1 igual a dois terços do mesmo comprimento em p2; p2 com o comprimento da paracrista igual a metade do comprimento da pós-metacrista comprimento mesio-distal de p2 e p3 iguais; altura da coroa de p3 um pouco maior que em p2; maior largura buco-lingual de p3 pouco maior que a mesma em p2 (fig. 39).

Molares
Molares Superiores - M3 com ectoestilo 1 reduzido, comparável ao ectoestilo j ; M3 com pequena redução da porção metastilar; M4 com ectoestilo 1 muito reduzido (fig. 40).
Molares inferiores – m4 com o comprimento mesio-distal pouco maior que m3; talonídeo em m4 mais estreito que nos demais (fig. 40).

 |
| Monodelphis emiliae Thomas 1912 |
Peramys emiliae Thomas (1912: 89)
[Didelphis (]Monodelphis[)] emiliae: Matschie, (1916: 271).
Monodelphis emiliae: Thomas (1920: 283).
P[eramys]. tricolor emiliae: A. Miranda Ribeiro (1936: 407).
P[eramys]. tricolor emilae: C. O. C. Vieira, (1950: 360).
Monodelphis tricolor emiliae: C. O. C. Viera (1955: 350).
Monodelphis (Monodelphis) touan emiliae: Cabrera (1958: 9) .
Monodelphis emiliae: Peterson & Pine (1982: 467).
Monodelphis emiliae: Pine & Handley, Jr. (1984: 239).
História taxonômica
Thomas (1912: 89) descreveu Peramys emiliae baseado em exemplares de Boim rio Tapajós, dois machos e uma fêmea. Comparou-a à Peramys scalops, salientando que a barriga é mais pálida e os dentes maiores. Em 1920, mais dois exemplares machos de Vila Braga, rio Tapajós, Pará, vêm complementar a descrição, observando que apresentavam uma curiosa "vinaceous coloration" por toda superfície ventral, já observada em um dos exemplares de Boim.
Miranda-Ribeiro (1936: 407) considerou P[eramys]. emiliae como uma subsepécie de P[eramys]. tricolor (Geoffroy) aplicando-lhe a designação, Peramys tricolor emiliae. Considerando inclusive P[eramys]. rubidus (Thomas) como outra subespécie, P[eramys]. tricolor rubidus. Quando Cabrera (1919: 43) colocou Didelphys brevicaudata (Thomas), na sinonímia de [Peramys] rubidus (Thomas), deveria ter colocado: (parte), pois citou:"Brasil oriental, Bahia". Provavelmente por não o ter feito isso, induziu Miranda-Ribeiro a pensar que Peramys rubidus estava sendo considerado como um sinônimo de Peramys tricolor (Geoffroy).
Muito pouco tem sido publicado sobre M. emiliae desde a descrição. No entanto, Pine & Handley Jr. (1984: 239) fizeram uma revisão da espécie utilizando material novo coletado no Brasil e no Peru. Ampliando dessa maneira sua área de distribuição geográfica até então restrita a margem esquerda do rio Tapajós. No Pará foi coletada em Cametá, margem esquerda do Tocantins, em Flexal (entre Itaituba e Jacareacanga) e no Km 217 da Santarém-Cuiabá (margem direita do rio Tapajós). Além disso tratam M. emiliae como uma espécie distinta, discordando de Miranda-Ribeiro, redescrevendo-a e comparando-a a outras espécies.
M. emiliae é sem dúvida uma espécie distinta, colocada aqui em um grupo próprio, postura que pode ser facilmente compreendida pela sua posição filogenética. A espécie é intermediária entre as espécies amazônicas do grupo M. brevicaudata e as do leste do Brasil com relação ao tamanho do corpo, dos dentes, forma da orelha e outros caracteres cranianos.(ver filogenia)
A pelagem é muito semelhante a de M. scalops, o que justifica a comparação de Thomas quando a descreveu, diferenciando-as pela coloração da barriga. O crânio é maior e mais curto que em M. scalops, particularmente o focinho, com bulas auditivas mais volumosas.
Descrição da pelagem
Coloração geral cinza e marrom avermelhada. Focinho, entre os olhos, na frente das orelhas, topo da cabeça e partes superiores dois membros anteriores marrons-alaranjados, mais alaranjado na frente das orelhas e no focinho, mais escuro no topo da cabeça; bochechas e lados do pescoço amarelados; nuca, dois terços anteriores do dorso e laterais do corpo cinza-grisalhos ou marrons-acinzentados-grisalhos, aristiformes com banda basal cinza e terminal preta ou sépia, setiformes com banda basal cinza, mediana preta ou sépia, subterminal branca-acinzentada e ponta preta ou sépia; terço posterior do dorso das laterais, partes superiores dos membros posteriores e parte dorsal, lateral e toda a base da cauda marrons-avermelhadas, pêlos aristiformes com banda basal cinza e terminal sépia ou marrom-alaranjada, pêlos setiformes com banda basal cinza, subterminal laranja e ponta sépia o cinza-grisalho e marrom-avermelhado do dorso se interpenetrando na área de contato; queixo, bordas da boca, garganta. peito, barriga e partes internas dos membros amarelo-ocre-acinzentados, pêlos de cor uniforme, mais claros no queixo, bordas da boca, garganta e membros, mais escuros na barriga; pêlos amarelos ventrais misturando-se com cinza das laterais; partes superiores das patas e partes inferiores da cauda com pêlos amarelado (fig. 41).
Variação: a coloração marrom-alaranjada da cabeça varia entre cinza-alaranjada-grisalha e marrom-alaranjada homogênea. Os dois, terços dorso-laterais variam entre cinza-grisalho e marrons-acinzentados grisalhos. Num exemplar examinado o dorso marrom levemente amarelado.
Comprimento dos pêlos:aristiformes, dorsais 8 mm, laterais 11 mm e ventrais 9 mm, setiformes, Dorsais e ventrais 6 mm e laterais 8 mm.
Medidas corporais: cabeça e tronco: 109-158 mm; cauda: 50-70 mm; 13-24 mm; orelha: 14-18 mm.
LINKs:
http://www.mammalogy.org/uploads/imagecache/library_image/library/1700.jpg
Espécimes examinados
Total: 3. BRASIL - Pará: Boim, 1 F (MG 539), Vila Braga, 1 M (MN 1299) - s/ loc 1 F (MN 1300)
http://www.mammalogy.org/uploads/imagecache/library_image/library/1700.jpg
Espécimes examinados
Total: 3. BRASIL - Pará: Boim, 1 F (MG 539), Vila Braga, 1 M (MN 1299) - s/ loc 1 F (MN 1300)

Monodelphis obscura, Gomes 1991
Descrição da pelagem
Coloração geral cinza, marrom-avermelhada e branca; focinho, parte superior da cabeça, ao redor das orelha, o terço posterior do dorso, partes superiores dos membros posteriores e cauda, marrons bem alaranjadas; nuca, dois primeiros terços do dorso, braços cinza-grisalhos, levemente amarronzados, aristiformes com banda basal cinza e terminal preta, setiformes com banda basal cinza, mediana preta, subterminal branca-acinzentada ponta preta; bochechas, queixo, bordas inferiores da boca, lados do pescoço, patas e brancos amarelados; garganta amarela-clara; peito, barriga e partes inferiores dos membros brancos levemente amarelados; pelos brancos desde a base; cauda com escamas visíveis apenas na superfície inferior (fig.41).
Comprimento dos pelos: aristiformes, dorsais e ventrais 8 mm, laterais 15 mm, setiformes, dorsais 5 mm, laterais 6 mm e ventrais 4 mm.
 |
Comparação entre M. emiliae e M. obscura características diagnósticas.
|
Baseada apenas em um exemplar sem crânio e sem localidade definida, essa espécie merece citação por apresentar algumas características bem distintas. Foi colocada no grupo M. emiliae por apresentar o mesmo padrão de pelagem, que no entanto nesta forma se apresenta mais evidente, com os pigmentos mais saturados. A cabeça, o terço posterior do dorso e as partes laterais das patas posteriores possuem pelos fortemente alaranjados comparados a M. emiliae. A parte superior do pescoço e os dois terços anteriores do dorso apresentam um cinza-grisalho bem definido, as bandas dos pelos aristiformes são mais escuras nesta espécie e as bandas subterminais dos setiformes são mais claras criando maior contraste. A superfície ventral é bem mais clara, quase branca. As orelhas são curtas e mais pilosas que em qualquer espécie do gênero, com pelos alaranjados. A cauda também é mais pilosa que nas outras espécies, com os pelos cobrindo toda a superfície dorsal (fig. 16). Diferentemente do que se observa no gênero, as escamas estão ausentes na superfície dorsal da cauda, ocorrendo apenas na parte ventral, nos dois terços terminais. O comprimento dos pelos dorsais é de cinco escamas aproximadamente (fig. 16). As almofadas plantares 1 da manus e do pes são alongadas e não ovaladas como nas outras espécies (figs. 14 e 15).
O exemplar examinado está depositado no Museu Paraense Emílio Goeldi. Eu o encontrei conservado em formol num recipiente grande, juntamente com espécies amazônicas. Como as coleções desse museu são praticamente restritas à amazônia, provavelmente trata-se de uma espécie dessa região, assim como M. emiliae.
Apesar de estar representada apenas por uma pele, sem crânio e sem localidade definida, esta é sem dúvida uma nova espécie por apresentar caracteres únicos dentro do gênero.
Espécime examinado
Total: 1 s / loc. sexo? (MG 11839).

 |
Monodelphis obscura, Estado do Acre - BRASIL
foto: Paulo S. Bernarde
|
 |
| Pelagem nos jovens de Monodelphis emiliae - exemplares de Juruena, MT (4) e Aripuanã, MT (3) |
 |
Variação na pelagem em adultos de M. emiliae - Aripuanã, MT e Santa Bárbara, RO e
à esquerda M. obscura - E. E. Antônio Mujica Nava, RO, BR com o colorido mais intenso e ventre quase branco. O tipo de M. obscura não tem localidade. Exemplares do MZUSP. |
 |
| Variação na pelagem em adultos de M. emiliae Aripuanã, MT e Santa Bárbara, RO e à esquerda M. obscura, E. E. Antônio Mujica Nava, RO, BR. |
 |
Variação na pelagem em adultos de M. emiliae
Aripuanã, MT e Santa Bárbara, RO e
à esquerda M. obscura, E. E. Antônio Mujica Nava, RO, BR. |
 |
| Monodelphis obscura, Pando, Bolivia Foto (c) Louise Emmons |
GRUPO Monodelphis americana
Características cranianas dos adultos
Maior comprimento do crânio 34,3 mm; forma do crânio (figs. 57-62); crista sagital presente ou ausente; perfil nasal entre quase reto e levemente convexo; forâmem infraorbital acima da região de contato entre P3 e M1; largura orbital entre um sexto e um sétimo do comprimento do crânio; jugal muito pouco dobrado na região orbital, ângulo interno variando normalmente entre 165º e 180º; processo pós-orbital do frontal, variando entre ausente e reduzido, afastado ou próximo da constrição pós-orbital; crista transversal do alisfenóide ausente, apenas uma leve ondulação em seu lugar; distância (drt) entre o foramen rotundum e o canal transverso aproximadamente igual a distância (dtc) entre o canal transverso e o forâmem da carótida; distância (dcc) entre os foramens das carótidas aproximadamente igual ou pouco maior que a distância entre cada um deles e o canal transverso adjacente; região posterior do palatino estreita, frontal muito próximo do forâmem orbitosfenóide; processo alisfenóide da bula ligado ao canal transverso na região mesial (pab), exceto em M. ihering; processo alisfenóide (pa) da bula terminando relativamente próximo ao processo timpânico do periótico; processo timpânico (pt) do periótico curto e arredondado, formando uma crista; perfil da borda inferior do ramo horizontal da mandíbula, entre levemente e quase reto; ângulo interno formado pelo processo coronóide e pelo ramo horizontal, entre 110º e 122º; processo coronóide estreito; inclinação interna do processo angular entre 110º e 130º; ramo ascendente da mandíbula com inclinação entre 15º e 30º (fig. 58).
Aspecto das bulas timpânicas no grupo M. americana:
 |
| Bulas timpânicas - note a redução dos processos anteriores nas formas do sul. Em M. iheringi até o processo timpânico do periótico reduziu-se. |
Diagnose: Identificamos o grupo pelas faixas dorsais ou pelas bulas
Variação da forma do crânio com a idade e entre as espécies.
Dentição
Pré-molares
superiores - diástema entre P1 e P2 nos adultos mais velhos, pode ser maior que o comprimento mesio-distal de P1 ou até um quarto desse comprimento; diástema entre P2 e P3 presente ou ausente; comprimento mesio-distal de P2 entre um terço e um quarto maior que P1 e entre pouco maior ou menor que P3; altura da coroa aproximadamente um terço menor que o comprimento mesio-distal em P2 (fig. 64).
inferiores - diástema entre p1 e p2 e entre p2 e p3; comprimento mesio-distal de p2 entre um terço e um quarto maior que p1 e pouco maior que p3; altura da coroa em p2 menor que o comprimento mesio-distal, em p3 aproximadamente igual (fig. 65).
Molares
superiores - comprimento mesio-distal de M1 pouco maior que em P3; M3 com ectoestilo l aproximadamente igual ao ectoestilo j; M4 com ectoestilo j geralmente muito reduzido.
 |
| Adicionar legenda |
inferiores - talonídio em m4 com a largura posterior aproximadamente igual a metade da mesma em m3.
Comparações cranianas: M. scalops, M. americana, fenótipo "rubida" (= M. americana), e M. iheringi (nas comparações utilize as figuras 49, 52, 53 e 57-67).
As espécies desse grupo apresentam um grau maior de variação que as espécies dos outros grupos. Como essa variação é gradativa, a comparação entre espécies morfologicamente próximas torna-se bastante útil na separação das espécies.
O fenótipo "rubida" (= M. americana) comparado com M. scalops apresenta crista sagital pouco mais pronunciada; processo pós-orbital do frontal mais evidente; crânio mais achatado dorsalmente; focinho mais longo; "bulas alisfenóides" relativamente maiores e mais fechadas; processo coronóide mais curto; dentes maiores; diástema entre P2 e P3, menos evidente; comprimento mesio-distal de P2 e P3, aproximadamente iguais (em M. scalops, P3‚ pouco menor que P2); diástema entre p2 e p3, menos evidente; comprimento mesio-distal de p2 e p3, aproximadamente iguais (em M. scalops, p3 é pouco menor que p2); p2 com coroa mais baixa; p3 com paracrista mais longa; ectoestilo j de M4 mais reduzido e o talonídeo de m4 mais alongado.
M. scalops comparada com a M. americana, apresenta a região dos frontais pouco mais alongada; anel timpânico pouco mais alongado junto ao processo timpânico do periótico; processo coronóide pouco mais largo; ângulo interno entre o processo coronóide e o ramo horizontal da mandíbula mais fechado; ramo horizontal da mandíbula com um estreitamento abaixo de p2 e p3; dentes menores; P2 com a paracrista menor que a pós-metacrista (em M. americana da Bahia a paracrista é aproximadamente igual a pós-metacrista); P3 com a coroa mais alta; P3 com a raiz anterior mais exposta que a posterior; p1 com a pós-metacrista uma vez e meia maior que a paracrista (em M. americana da Bahia, a pós-metacrista é duas vezes maior que a paracrista); p3 mais alto, p3 com a pós-metacrista duas vezes maior que a paracrista (em M. americanada Bahia, a pós-metacrista é uma vez e meia maior que a paracrista); ectoestilo j mais evidente em M4; talonídeo mais largo em m4.
M. americana da Bahia, comparada com M. americana do Pará, apresenta o crânio e o palato mais alongado; crista sagital mal formada (naqueles do Pará é evidente nos adultos); ramo horizontal da mandíbula mais largo abaixo de m4.
M. americana do Pará, comparada a M. americana do sul, apresenta o palato mais largo, focinho mais robusto, nasais mais largos, distância entre o foramen rotundum e o canal transverso um pouco maior que a distância entre o canal transverso e o forâmem da carótida adjacente; Naqueles do Pará a distância entre o foramen rotundum e o canal transverso é aproximadamente igual a distância entre o canal transverso e o forâmem da carótida adjacente; ramo horizontal da mandíbula um pouco mais alargado na região de m1 e m2; processo coronóide ligeiramente mais largo; dentes menores; P1, P2 e P3 com coroas um pouco mais altas; M4 pouco mais largo no sentido antero-posterior do palato; m4 mais longo; cíngulo bucal do talonídeo de m4 um pouco mais estreito.
M. iheringi comparada com M. americana do sul, apresenta crânio menor; crista sagital levemente sugerida em M. americana (em M. iheringi não forma-se crista sagital); processo anterior da bula ausente em M. iheringi [Diagnose]; perfil da região frontal convexo, reto em M. iheringi; focinho mais longo; processo coronóide mais longo; dentes maiores; incisivos inferiores pouco mais proodontes; P1 e p1 com o comprimento mesio-distal menor; p2 e p3 com a coroa mais alta; m4 com a largura buco-lingual um pouco maior.
 |
| Monodelphis americana Muller 1776 |
Mus araneus: Marcgrave (1648: 229).
La musaraigne du Brèsil: Buffon (1767: 160).
Brasilian Shrew: Pennant (1771: 309).
Sorex americanus Muller (1776: 36).
Sorex brasiliensis Erxleben (1777: 127). Schreber (1778: 577). Boddaert (1785: 125). Gmelin (1788: 115). Kerr (1792: 206).
Sorex brasiliensis: Zimmermann (1780:386). Shaw (1800: 535).
Didelphys tristriata Illiger (1815: 107). Kuhl (1820: 63). Fischer (1829: 269). Waterhouse (1841: 107). Schinz (1844: 511). Waterhouse (1846: 518). Wagner (1848: 52).
Didelphis tristriata Schinz (1821: 252).
Didelphys trilineata Lund (1841: 237).
Peramys triistriata: Lesson (1842: 187).
Microdelphys tristriata: Burmeister (1856: 84).
Didelphys (Peramys) americana: Thomas (1888b: 363).
Peramys americanus: Cabrera (1919: 42). Miranda-Ribeiro (1936: 414).
Peramys iheringii: Miranda-Ribeiro (1936: 415) parte.
Monodelphis americana: Cabrera & Yepes (1940: 30). Emmons (1990: 30) parte.
Monodelphis americanus: Vieira (1949: 358 parte. Vieira (1955: 349) parte. Hershkovitz (1959: 338).
Monodelphis (Monodelphis) americana americana: Cabrera (1958: 6) parte.
Monodelphis rubida: Emmons (1990: 30) parte.
Peramys tristriata: Goeldi & Hagmann (1904: 100)
Minuania umbristriata: Miranda-Ribeiro (1936: 422).
M[inuania]. goyana: Miranda-Ribeiro (1936: 419).
Minuania umbristriata: Cabrera & Yepes (1940: 33).
Monodelphis umbristriatus: Vieira (1955: 350).
Monodelphis (Monodelphis) americana americana: Cabrera, (1958: 6) parte.
Monodelphis rubida: Emmons (1990: 30) parte.
Localidade tipo: "Brasilien," restrita para Brasil, Pernambuco, por Cabrera (1958:7).
[VOU RETIFICAR ISSO AQUI, ESTOU REVISANDO TODO TEXTO on line ]
História taxonômica
Esta foi a primeira espécie do gênero a ser conhecida pela ciência. Descrita de uma forma sucinta por Marcgrave (1648: 229) como Mus araneus (rato aranha), não foi reconhecida como um marsupial antes de 1815. Em sua descrição Marcgrave comparou-a a um rato. "Mus araneus com a forma do rato. O comprimento do corpo, da borda da boca até a extremidade da cauda, tem cerca de cinco polegadas; a cauda tem cerca de duas (polegadas). Possui o focinho pontudo, e os dentes agudíssimos: escroto com os testículos pendentes entre as pernas traseiras. A coloração é escura, mas da parte posterior da cabeça até a cauda, tem três linhas negras grossas longitudinais. Brinca com o gato, não tem medo dele, nem ele o caça".
Buffon (1767:160) descreveu-a como "La musaraigne du Brézil", Pennant (1771:309) citou-a como "Brazilian Shrew", ambos baseados em Marcgrave,
Porém, Muller (1776:36) foi o primeiro a aplicar-lhe a nomeação binomial lineana válida: Sorex americanus. Colocando-a no mesmo gênero do musaranho europeu. Em 1777, um ano depois, Erxleben descreveu-a como Sorex braziliensis. Ambos se basearam em Buffon.
Illiger (1815: 107) foi o primeiro a denominá-la como um marsupial, tratando-a como [Didelphis] Tristriata. Muito provavelmente baseado nas observações que Azara havia feito sobre o musaranho do Brasil de Buffon.
Miranda-Ribeiro (1936: 422) descreveu-a como Minuania umbristriata baseado em dois exemplares provenientes de Goiás, um deles, um adulto com eritrismo. Localidade apontada mais precisamente como: "Veadeiros, Goiás" por Avila-Pires, em 1968.
Vieira (1955: 350) tratou-a como Monodelphis umbristriatus, errando a concordância.
Pine (1976:785) considerou Monodelphis umbristriata (de Miranda-Ribeiro) como uma espécie válida, e diferente de M. americana, baseado em um exemplar macho (do National Museum of Natural History) de Passos, Minas Gerais. Ele comparou a descrição de Miranda-Ribeiro com o exemplar citado, que ele acreditou ser este o terceiro exemplar conhecido até então. Considerou-a como membro do subgênero Minuania e simpátrica com M. americana. Pine cita as diferenças entre dois exemplares de Passos, MG. As diferenças apontadas por Pine entre os dois exemplares de Passos do USNM, devem ser relacionadas a idade ou ao sexo. Pine não cita o sexo desse segundo exemplar, mas as diferenças usadas na distinção dessas espécies são aquelas observadas entre os machos e as fêmeas da espécie M. americana dessas regiões.
Vieira (1955: 350) tratou-a como Monodelphis umbristriatus, errando a concordância.
Pine (1976:785) considerou Monodelphis umbristriata (de Miranda-Ribeiro) como uma espécie válida, e diferente de M. americana, baseado em um exemplar macho (do National Museum of Natural History) de Passos, Minas Gerais. Ele comparou a descrição de Miranda-Ribeiro com o exemplar citado, que ele acreditou ser este o terceiro exemplar conhecido até então. Considerou-a como membro do subgênero Minuania e simpátrica com M. americana. Pine cita as diferenças entre dois exemplares de Passos, MG. As diferenças apontadas por Pine entre os dois exemplares de Passos do USNM, devem ser relacionadas a idade ou ao sexo. Pine não cita o sexo desse segundo exemplar, mas as diferenças usadas na distinção dessas espécies são aquelas observadas entre os machos e as fêmeas da espécie M. americana dessas regiões.
Durante o estudo que realizei com o grupo, observei dentro da variação de colorido da pelagem de M. americana um fenômeno bastante interessante. Alguns exemplares machos de idade avançada apresentam uma ausência de pigmento preto que pode ser total ou parcial. Nesses exemplares os pelos são geralmente rígidos ao tato e curtos, permanecendo neles apenas o pigmento vermelho ou alaranjado. Aparentemente os machos velhos em uma certa área de distribuição da forma meridional podem desenvolver uma coloração de pelagem avermelhada. O pigmento preto desaparece ou é substituído por pigmento vermelho, e as três faixas longitudinais pretas tornam-se marrom-avermelhadas.
Entre o material que examinei no Museu Nacional, Rio de Janeiro, havia dois machos e três fêmeas, da localidade de Passos, em Minas Gerais, além dos dois exemplares de "Minuania umbristriata" (de Miranda-Ribeiro). De acordo com as medidas realizadas e as características observadas, os exemplares de Passos não correspondem exatamente àqueles utilizados na descrição de M. umbristriata. Um dos machos de Passos apresentava uma tonalidade avermelhada geral provocada pela ausência do pigmento preto e o outro apresentava uma perda de pigmento apenas na região anterior do corpo. Esse fenômeno, citado acima, foi também observado em exemplares de Mariana, MG, e de Piracicaba, Piquete e São Paulo, SP (todos do MZUSP). Considero M. umbristriata, como sinônimo júnior de M. americana.
Esse avermelhamento dos machos velhos é aqui tratado como eritrismo. O exame destas peles permite-nos observar apenas um leve traço das listas negras dorsais tão características desta espécie. As listas aparecem avermelhadas ou marrom avermelhadas, sempre muito pouco visíveis. Outros indivíduos podem ser bastante escuros apresentando a lista central mais evidente que as laterais. Esse eritrismo foi constatado no exemplar adulto de M. umbristriata, usado na descrição de Alipio de Miranda-Ribeiro, mas não no lectótipo, um imaturo designado por Paulo de Miranda-Ribeiro. Os exemplares de Minas Gerais e São Paulo compartilham essa característica com o exemplar adulto de M. umbristriata, mas são menores e correspondem mais, em outras características, a forma meridional do grupo M. americana, ou seja, M. iheringi.
O exemplar adulto de M. umbristriata, quanto ao tamanho e ao formato do crânio, corresponde mais aos exemplares do Estado do Pará. O crânio é maior, com os nasais e toda a parte posterior do focinho proporcionalmente mais larga, dentes maiores e crista sagital mais desenvolvida que nos exemplares de M. americana de Minas Gerais. O eritrismo tem uma distribuição independente dos caracteres cranianos.
Em sua descrição Miranda-Ribeiro baseou-se principalmente na pelagem do adulto para caracterizar a espécie, mas cita também "a presença do processo supraorbitário e a conformação dos nasais, tamanho dos caninos, largura zigomática, p3 sendo maior e outros aspectos do craneo". Uma das características usadas por Miranda-Ribeiro na sua definição do gênero Minuania, foi "a presença de crista sagital procedendo de perto dos processos orbitaes". O exame do material de Minas Gerais de M. americana mostrou que a crista sagital só aparece nos machos muito velhos (fig.63), enquanto em nos do Pará já está bem desenvolvida nos adultos mais jovens (fig.63).
O exemplar adulto de M. umbristriata, assim como os exemplares do Pará, apresenta a cabeça avermelhada sem a lista negra central, enquanto que nos exemplares de Minas Gerais a lista negra central está presente na parte anterior do focinho e pode aparecer enfraquecida no resto da cabeça. Observa-se ainda nos exemplares do Pará apenas a lista central no traseiro, que é mais avermelhado e a parte ventral mais amarelada enquanto no sul as três listas chegam até a base da cauda e a barriga é acinzentada com muito pouco amarelo. Essas duas formas parecem representem apenas um gradiente geográfico com ausência de coletas na sua zona intermediária.
O mapa da fig. 69 mostra a área onde ocorre o eritrismo. As localidades citadas estão plotadas ao redor do que considero a linha de dispersão da espécie. Ao norte a localidade tipo de M. umbristriata. As localidades onde o eritrismo foi detectado, estão aparentemente associadas a regiões onde exite uma mistura de mata com cerrado ou campos abertos.
Em meu tranbalho de 1991, como já citei acima, considerei M. umbristriata como espécie válida. No entanto nesta atualização examinei os exemplares do MZUSP que conta atualmente com material novo coletado na última década. O material coletado no Ceará foi muito importante na minha decisão de considerar M. umbristriata como parte da variação geográfica de M. americana. Os exemplares do Ceará possuem o traseiro avermelhado de maneira semelhante aos do Pará, no entanto parecem intermediarios entre estes e os de Pernanbuco e Paraíba. Tal semelhança mostra que existe uma gradação entre a forma do nordeste, de onde vem o tipo de M. americana, e as formas do Pará e do Brasil central de onde vem os exemplares de M. umbristriata. Sendo assim, não é possível considerar M. umbristriata com uma espécie distinta, pois não existe diagnose possível para caracterizar-la. A coloração de listras indistintas que deu o nome a espécie não representa mais do que uma coloração ligada ao sexo e a idade, assim como as características cranianas.
Ronald Pine & C. Handley, Jr. (1984: 242) incluiram [Monodelphis] umbristrita em M. rubida (de Thomas) e foram muito provavelmente seguidos por Emmons (1990: 30), que considerou Goiás, Minas Gerais e São Paulo como parte da distribuição geográfica de M. rubida, citando também as três faixas dorsais marrons-escuras, como aquelas observadas no exemplar adulto de Monodelphis umbristriata.
Os exemplares (que examinei) de Monodelphis rubida (de Jaguaquara, na Bahia) correspondem admiravelmente a descrição de Thomas (1899: 155). Porém, Thomas não citou nada sobre faixas dorsais na coloração da pelagem, como nos exemplares que Miranda-Ribeiro usou na descrição de M. umbristriata, os quais não correspondem exatamente à M. rubida (de Thomas). Ao que tudo indica, as faixas podem desaparecer por completo, em indivíduos muito velhos, como parecem ser os de Jaguaquara.
Lemos et al. (2000 :329) chamam à atenção para essa confusão existente entre M. rubida e M. umbristriata. Inclusive eles usaram a minha tese de mestrado de 1991 que já apontava tal confusão. Nesse trabalho, eles reconheceram, como eu em 1991, o status de M. umbristriata como espécie válida. No entanto não dispunham de material suficiente para detectar a existência do eritrismo nas populacões do sul. Consideraram "The most notable difference is the faint longitudinal stripes on the dorsum of M. umbristriata" como característica principal da espécie. Na comparação com M. americana utilizaram um adulto jovem que denominaram como "dental age seven" que na verdade não possui idade comparável. Podemos perceber que na fig.2 da pág. 333 que o exemplar de M. americana (MN 11179) possui a região posterior do palato ainda pouco projetada para trás e na fig.3 da pág. 334 o P3 não possui as raízes expostas como no exemplar MN 46570. Nos adultos velhos (como no caso do exemplar MN 46570) essas características são diagnósticas da idade avançada assim como a presença de crista sagital. Os processos pós-orbitais desenvolvidos realmente chamam à atenção, e de fato não encontram paralelo dentro do gênero. Mesmo assim, não são suficientes por si só para caracterizar M. umbristriata, e podem apenas ser considrados como uma possível variação local ou devido a idade muito avançada do exemplar. Tal característica não deve aparecer nos exemplares mais jovens ou nas fêmeas, não podendo portanto ser utilizada como uma característica diagnóstica.
Descrição da pelagem dos exemplares da Bahia
Parte superior da cabeça, lados do focinho, bochechas, ao redor das orelhas, partes superior e laterais do pescoço, dois terços anteriores do dorso e laterais, marrom amarelado, mais vivo ao redor das orelhas e bochechas, pêlos aristiformes com banda basal cinza e terminal sépia, pêlos setiformes dorsais com banda basal cinza, terminal sépia nas faixas e pêlos setiformes dorsais e laterais com banda basal cinza, subterminal amarela e ponta preta; terço posterior do dorso, traseiro e partes superiores dos membros, marrom avermelhado, mais atenuado nos ombros anteriores; três faixas pretas ao longo do dorso; faixa central da ponta do focinho até a base da cauda, as laterais dos membros a base da cauda; queixo e bordas inferiores da boca, amarelo claro rosado; garganta, peito, barriga e partes internas dos ombros, ocre acinzentado, pêlos aristiformes com banda basal cinza, subterminal sépia e ponta amarelo claro, pêlos setiformes com banda basal cinza e terminal amarelo claro; partes superiores das patas, marrom alaranjado, mais claro nas patas traseiras; patas posteriores inferiormente, negras, dedos com escamas; cauda com pêlos iguais ao corpo apenas na base, o restante com escamas parcialmente cobertas por pequenos pêlos, preto em cima, marrom por baixo (fig. 68).
Variação: O jovem MN 11492 apresenta as partes inferiores do corpo mais acinzentadas que os adultos, as bandas amarelas terminais dos pêlos são mais curtas. Alguns indivíduos apresentam as bandas amarelas dos pêlos ventrais mais largas, resultando um ruivo mais acinzentado. O MN 20976 apresenta um avermelhado geral por toda pelagem, lados do focinho, ao redor das orelhas, marrom alaranjado; bochechas alaranjadas; as laterais avermelhadas. A faixa central pode estar interrompida entre os olhos ou diluída na cor da cabeça.
Comprimento dos pêlos: aristiformes, dorsais 7 mm, laterais 8 mm, ventrais 6 mm; setiformes, dorsais 4 mm, laterais 4 mm e ventrais 4 mm.

Descrição da pelagem do fenótipo meridional
Coloração geral marrom avermelhado ou acinzentado com estrias escuras no dorso. Parte superior da cabeça, lados do focinho, bochechas, pescoço, dorsal e lateralmente, dois terços anteriores do dorso e laterais, marrom acinzentado ou marrom avermelhado, mais amarelado ao redor das orelhas, pêlos aristiformes dorsais e laterais com banda basal cinza e terminal sépia; pêlos setiformes dorsais, com banda basal cinza e terminal sépia nas listras e com banda basal cinza, mediana sépia, subterminal branco amarelado e ponta preta, no resto; terço posterior do corpo, partes superiores dos membros marrom avermelhado ou levemente avermelhado, pêlos aristiformes com banda subterminal alaranjada; três faixas pretas ou marrom ao longo do dorso; a faixa central estendendo-se da ponta do focinho até a base da cauda, difusa ou ausente entre os olhos, mais nítida a partir do topo da cabeça, faixas laterais desde os ombros até a base da cauda; queixo, bordas inferiores da boca, garganta, amarelo claro esbranquiçado, pêlos sem banda basal cinza; pescoço inferiormente, peito, barriga e partes internas dos membros, cinza amarelado, pêlos aristiformes e setiformes com banda basal cinza e ponta branca amarelada; partes superiores das patas, marrom claro, mais amarelado nas patas traseiras; plantas das patas posteriores levemente enegrecidas, anteriores mais claras; cauda com pêlos iguais aos do corpo apenas na base, o restante com escamas parcialmente cobertas por pequenos pêlos, marrom em cima e marrom amarelado em baixo (fig. 68).
Variação: a coloração dorsal de fundo pode ser mais acinzentada ou mais amarelada, dependendo das bandas subterminais dos pêlos setiformes. O marrom avermelhado do traseiro pode ser muito claro ou não existir. Os exemplares subadultos de Além Paraíba, MG, possuem a coloração de fundo dorso-lateral quase uniforme, apenas levemente alaranjada no traseiro, partes superiores dos membros e na frente das orelhas. Dois machos adultos de Passos, MG, MN 20971 e 20972, possuem a coloração dorso lateral quase uniformemente ruiva, marrom avermelhada no traseiro, membros e topo da cabeça. As faixas dorsais variam entre preto, sépia e marrom avermelhado, no MN 20971, são marrom avermelhadas no primeiro terço do dorso e negras nos dois terços finais. Ventralmente de acordo com as bandas, basal cinza e amarelo claro dos pêlos, o cinza pode ser esbranquiçado, mais escuro ou amarelado. Aparentemente os machos velhos em uma certa área de distribuição da forma meridional podem desenvolver uma coloração de pelagem avermelhada.O pigmento preto desaparece ou é substituído por pigmento vermelho, as três faixas longitudinais pretas apresentam-se marrom avermelhadas. Observei exemplares assim descritos em Piracicaba, Piquete, São Paulo, Mariana e Passos. Poderíamos incluir nessa lista os exemplres intermediários de Além Paraíba e o holótipo de M. umbristriata.
Comprimento dos pêlos: aristiformes, dorsais e laterais 7 mm e ventrais 5 mm; setiformes, dorsais e laterais 4 mm e ventrais 3 mm.
Medidas corporais: cabeça e tronco: 95-125 mm; cauda: 43-63 mm; pé: 14-21 mm; orelha: 8-16 mm.
Parte superior da cabeça, lados do focinho, bochechas, ao redor das orelhas, marrom levemente alaranjado, mais alaranjado ao redor das orelhas e nas bochechas, mais escuro no topo da cabeça; parte superior do focinho, enegrecido, as vezes com pequeno segmento preto longitudinal; pescoço, dois terços anteriores do dorso e laterais, marrom levemente amarelado, pelos aristiformes, dorsais e laterais com banda basal cinza e terminal sépia, pelos setiformes dorsais com banda basal cinza e terminal sépia nas faixas e com banda basal cinza, mediana sépia, subterminal amarelo claro e ponta sépia, no resto, pelos setiformes, laterais com banda basal cinza, mediana sépia, subterminal amarelo claro e ponta sépia; o terço posterior do dorso, traseiro e partes superiores dos membros, marrom levemente avermelhado; três faixas pretas ao longo do dorso; faixa central do topo da cabeça a base da cauda, mais difusa na região marrom avermelhada do traseiro; faixas laterais mais curtos, evidentes apenas no terço central do dorso, diluindo-se anteriormente até os ombros e quase imperceptíveis no terço posterior do dorso; queixo, bordas inferiores da boca e garganta amarelo claro rosado, pelos sem banda basal cinza; pescoço inferiormente, peito, barriga e partes internas dos membros, ocre acinzentado, pelos aristiformes, com banda basal cinza, subterminal sépia e ponta amarelo claro, pelos setiformes, com banda basal cinza e terminal amarelo claro; parte superior das patas, marrom claro mais claro nos pés; dedos com escamas; cauda com escamas visíveis e pelos na base, exceto na base inferiormente, marrom escuro superiormente e mais claro inferiormente (fig. 68).
Variação: Alguns indivíduos possuem toda a pelagem mais avermelhada e as faixas dorsais menos nítidas. As bandas amarelas dos pêlos ventrais podem variar na largura, originando um ocre muito pouco acinzentado ou um cinza amarelado. Esta forma foi citada pela primeira vez por Goeldi & Hagmann (1904: 100) como Peramys tristriata Illiger."...adulto nas vizinhanças do Pará. O colorido geral‚ consideravelmente mais escuro que o da estampa no. 35 da recente obra "Hand-book of the Marsupialia".
Comprimento dos pêlos: aristiformes, dorsais 6 mm, laterais e ventrais 7 mm; setiformes, dorsais 4 mm, laterais 4 mm, ventrais 3 mm.
Medidas corporais: cabeça e tronco: 59-135 mm; cauda: 38-61 mm; pé: 14-22 mm; orelha: 12-16 mm.
 |
Machos de M. americana com eritrismo, ocorre no Brasil central. Minas Gerais, São Paulo e Espírito Santo.
Corresponde ao avermelhamento de "rubida". Veja o holótipo de M. rubida acima à direita. |
LINKS:
Total: 58. BRASIL - Pará: Mosqueiro, 1 M (MG 3543); Belém, 5 M (MZUSP 11698, MG 318, 327, 1975, 2573), 2 F (MZUSP 9930, MG 2574), 2 ? (MG s/n, MZUSP 8237); 1 M (MN 1304), 2 F (MN 1303, 1305). - Goiás: Veadeiros, 1 M (MN 1313), 1 ? (MN 1314). - Brasília: 1 F (UFPB 2833). - s/loc. 2 M (MZUSP 9415, 10206), 2 F (MN 315, 316).- Paraíba: João Pessoa, 1 M (APO 406), 1 F (UFPB 413); -Pernambuco: Caruarú, 1 M (MN 24544). - Bahia: Ilhéus, 6 M (MN 11075, 11179, 11180, 11485, 11492, 20976); Ilhéus ?, 1 M (MN 11524), 2 F (MN 11483, 11498). - Minas Gerais: Além Paraíba, 2 M (MN 7569, 7571), 1 F (MN 7568); Mariana, 1 M (MZUSP 2193, 1 F (MZUSP 3418); Passos, 2 M (MN 20971, 20972), 3 F (MN 11728, 20973, 20974); Porto Novo, 1 F (MN 7312). - Espírito Santo: Santana, 1 M (MN 1307); - Rio de Janeiro: Itatiaia, 1 F (MZUSP 430); Teresópolis, 1 F (MN 7250). - São Paulo: Paraibuna, 1F (MN 10988); Perus, 1F (MZUSP 11); Piquete, 2 ? (MZUSP 118, 1520); Piracicaba, 4 M (MZUSP 1516, 1518, 1519(a), 1519(b)), 1 F (MZUSP 1523); São Paulo, 2 M (MZUSP 1185, CNFG 1), 1 F (CNFG 2).
 |
M. americana (descrição do fenótipo histórico "rubida")
|
Didelphys (Peramys) brevicaudata: Thomas (1888b: 356) parte.
Peramys rubidus: Thomas (1899:155). Cabrera (1919: 43).
P[eramys]. tricolor rubidus: Miranda – Ribeiro (1936: 408).
Monodelphis tricolor rubidus: Vieira (1955: 350).
Monodelphis (Monodelphis) touan rubidus: Cabrera (1958: 10).
[Monodelphis] rubida: Pine & Handley, Jr. (1984: 242) parte.
M. rubida Lemos et. al. (2000 :329).
Localidade tipo: Brazil, "Bahia."
Historia taxonômica
Localidade tipo: Brazil, "Bahia."
Historia taxonômica
Thomas (1888b: 356) no seu "Catalogue of Marsupials", descreveu Didelphys (Peramys) brevicaudata com base em seis exemplares. Um da Bahia (macho), outro que foi usado por Waterhouse na descrição de Didelphys hunteri e mais quatro sem localidades mencionadas. De acordo com a descrição apresentada, pelo menos duas diferentes espécies estavam englobadas nesse material. O holótipo de D. hunteri, corresponde à Monodelphis brevicaudata Erxleben. Daqueles quatro, pelo menos dois exemplares, sem localidade, representavam M. brevicaudata, a julgar pelo comprimento da série molar M1-M2: 6,2 e 6,8 mm, e pelos tipos de padrões de colorido descritos. O exemplar da Bahia foi mais tarde descrito por Thomas (1899: 155) como "Peramys rubidus", justificando no final da descrição que "os caracteres relacionados à cor naquela época não pareciam tão importantes quanto provaram ser". Na descrição comparou a nova espécie com "P[eramys]. brevicaudatus". Considerando o tamanho, as proporções e os caracteres cranianos como em "P. brevicaudatus", embora o focinho seja um pouco mais afilado. Cor: ruiva-acastanhada uniforme, por todo o dorso e lados; na cabeça um pouco mais vivo e no traseiro um pouco mais escuro. Barriga, cinza amarelado opaco, pêlos marrons acinzentados na base e amarelos claros opacos nas pontas. O lado externo dos membros, as partes dorsais das mãos e pés ruivo opaco. Cauda toda ruiva. Medidas: corpo: 160,0; cauda: 64,5; orelha: 13,0; crânio com largura zigomática: 19,4; nasais: 18,0 x 5,7; largura inter-orbital: 6,6; comprimento M1-M3: 5,9 (todas em mm).
A descrição de Thomas corresponde perfeitamente aos exemplares MN 24550, 24551, do Museu Nacional coletados em Jaguaquara, Bahia. Portanto, a única localidade precisa para esta espécie (aqui considerada como uma forma geográfica).
Emmons (1990: 30) considerou Goiás, Minas Gerais e São Paulo como parte da distribuição geográfica de M. rubida, certamente examinou machos de M. americana com eritrismo, essa identificação provavelmente originou-se na referência de Pine & Handley, Jr. (1984: 242) incluíndo [Monodelphis] umbristriata (= M. americana) em M. rubida (Thomas).
Lemos et al. (2000: 329) chamam à atenção para a confusão existente entre M. rubida e M. umbristriata. Nesse trabalho usaram a minha tese de mestrado de 1991, que já apontava tal confusão. Atualmente mudei de opinião sobre M. umbristriata, novo material coletado mostrou que não é possível separa-la de M. americana, as diferenças existentes entre ambas são simplesmente variações geográficas (ver detalhes no grupo M. americana).
O fato de M. rubida ser conhecida apenas por três exemplares machos, e a sua proximidade morfológica e evolutiva com M. americana, está certamente relacionada ao eritrismo observado no Brasil central (dimorfismo sexual). M. rubida atualmente já é amplamente reconhecida com uma forma de eritrismo, uma manifestação geográfica de M. americana (tanto o tipo, como os exemplares de Jaguaquara do MNRJ, são machos).
Descrição da pelagem do fenótipo com eritrismo conhecido como "rubida"
Coloração geral marrom-alaranjado. Focinho, parte superior da cabeça, dorso, laterais e partes superiores dos membros marrons alaranjados, mais vivo nos ombros e membros anteriores, mais opaco no traseiro e membros posteriores, pêlos aristiformes dorsais com banda basal cinza, subterminal preta e terminal laranja, ou com banda basal cinza e terminal preta, ou com banda basal cinza, mediana preta, subterminal laranja e ponta preta; pêlos setiformes dorsais com banda basal cinza, subterminal laranja e ponta preta; pêlos aristiformes laterais com banda basal cinza, subterminal preta e ponta laranja, pêlos setiformes laterais com banda basal cinza, subterminal laranja, bem larga e terminal preta, pequena; bochechas, queixo e bordas inferiores da boca, amarelo-claro esbranquiçado, pêlos sem banda basal cinza; parte inferior do pescoço, peito, barriga e partes internas dos membros, cinzas-amarelados, pêlos com banda basal cinza e terminal amarelo-claro; patas alaranjadas; cauda com pêlos iguais ao corpo apenas na base, o restante com escamas cobertas por pêlos pequenos, dorsalmente, marrom-claro, ventralmente marrom-amarelada-clara (fig. 54).
Comprimento dos pelos: aristiformes, dorsais 10 mm, laterais 11 mm, ventrais 11 mm: setiformes, dorsais 6 mm, laterais 7 mm e ventrais 4 mm.
Medidas corporais: cabeça e tronco: 126-160 mm; cauda: 65-70 mm; pé: 18-21 mm; orelha: 13-18 mm.



Esses dois exemplares de M. americana do Espírito Santo apresentam uma semelhança muito grande com as peles que examinei de M. "rubida", se não fosse pela presença das faixas alaranjadas o resto da pelagem é idêntico àquelas. O crânio com processos pós-orbitais também concorda. Os processos pós-orbitais aumentam com a idade avançada e aparentemente são mais desenvolvidos nessas populações de Goiás, MG, ES e Bahia. O que explicaria as formas que originaram M. umbristriata e M. rubida. Em São Paulo M. americana apresenta tamanho de crânio menor sem pocessos pós-orbitais, forma-se uma crista sagital muito pouco pronunciada e sem dúvida é uma característica gradual.
 |
| Monodelphis americana, Diamantina, Minas Gerais, Brasil Um macho com eritrismo - foto: Marco Aurélio Pacheco. |
 |
| Monodelphis americana by Rodrigo Conte Jardim Botânico de Brasilia, Brasil |
 |
| Monodelphis americana, Floresta Nacional de Ipanema em Sorocaba, São Paulo - BRASIL |
MAPA DE DISTRIBUIÇÃO
 |
No mapa temos a indicação da única localidade precisa de M. "rubida" (=M. americana) compreendida numa visão restrita ou seja a descrição original sem interpretações. M. rubida faz parte do fenômeno do eritrismo em M. americana, representa simplesmente uma forma local, uma resposta ao limite de aridez que existe na região indicada para a espécie (Caatinga). A inexistência de simpatria entre M. "rubida" e M. americana corrobora a ideia de variação local.
|
 |
| Monodelphis iheringi Thomas 1888 |
Didelphis (Peramys) iheringi: Thomas (1888a: 159). Thomas (1888b: 364)
[Peramys] americanus: Cabrera (1919: 42) parte.
Peramys iheringii: Miranda-Ribeiro (1936: 415) parte.
Monodelphis americana: Cabrera & Yepes (1940: 30) parte.
Monodelphis iheringii: Vieira (1949: 359) parte. Vieira (1955: 349) parte.
Monodelphis americanus: Vieira (1949: 358) parte. Vieira (1955: 359).
Monodelphis (Monodelphis) america iheringi: Cabrera (1958: 7)
Monodelphis iheringi: Pine (1977: 235). Emmons (1990: 30).
Localidade tipo:"Rio Grande do Sul;" identificada como Brasil, Rio Grande do Sul, Taquara, by Thomas (1888).
História taxonômica
Esta espécie foi descrita por Thomas (1888a: 159) baseada em um macho do "Rio Grande do Sul" como "Didelphys (Peramys) iheringi". Thomas comparou-a com M. americana (Muller), citando que as cores e as proporções são exatamente como nessa espécie, mas com a metade do tamanho desta. Em descrições posteriores Thomas (1888b: 364) adicionou que Monodelphis iheringi caracteriza-se por um crânio achatado e forneceu como localidade típica "Taquara, Rio Grande do Sul".
Miranda-Ribeiro (1936: 415) listou um exemplar desta espécie de "Japaranã (Sant'Anna), E[stado]. do Espírito Santo". Examinei esse exemplar no Museu Nacional (RJ) e identifiquei-o como M. americana (Muller).
Vieira (1949: 375 e 1955: 349) listou um exemplar de "Chaves, Estado do Espírito Santo". Também examinei esse exemplar no Museu de Zoologia da USP e identifiquei-o como M. scalops (Thomas).
Cabrera (1958: 7) tratou M. iheringi como uma subespécie de M. americana (Muller). Pine (1977: 235) publicou uma nota reconhecendo Monodelphis iheringi (Thomas) com uma espécie válida, diferente de M. americana. Aqui sigo o mesmo caminho reconhecendo a espécie. M. iheringi, que possui pelagem idêntica às populações de M. americana do sudeste do Brasil ou mais clara, no entanto, possui tamanho menor e características cranianas próprias, sendo diagnosticáveis pela comparação das bulas (ver fig. 23)
A fig. 70 mostra a relação de tamanho existente entre os molares superiores de M. iheringi e M. americana. Podemos notar que as dimensões das amostras de M. americana do sudeste de sua distribuição (do Espírito Santo até Santa Catarina), estão mais próximas das de M. iheringi que as demais.
Descrição da pelagem
Coloração geral marrom acinzentado com estrias no dorso. Parte superior e laterais da cabeça e dois terços do corpo, marrom acinzentados, pelos aristiformes dorsais e laterais com banda basal cinza e terminal sépia, setiformes dorsais com banda basal cinza e terminal sépia nas faixas e com banda basal cinza, subterminal branco amarelado e ponta sépia no resto, setiformes laterais com banda basal cinza, subterminal amarelo claro e ponta sépia; ao redor das orelhas, terço posterior do corpo e partes superiores dos membros, marrom avermelhado; três faixas pretas ao longo do dorso; a central estendendo-se da ponta do focinho a base da cauda, difusa entre os olhos, faixas laterais a partir dos ombros a base da cauda; queixo bordas da boca e garganta amarelo claro esbranquiçado, pelos sem banda basal cinza; parte inferior do pescoço, peito, barriga e partes internas dos membros, cinza amarelado, pelos com banda basal cinza e terminal branco amarelado; partes superiores das patas, marrom alaranjado, mais vivo nas patas posteriores; plantas das patas posteriores levemente enegrecidas, anteriores mais claras; cauda com pelos iguais aos do corpo apenas na base, o restante com escamas parcialmente cobertas por pelos, dorsalmente marrom, ventralmente marrom amarelado (fig. 68).
Variação: dos sete exemplares examinados, MN 1308 e MN 1309, da Restinga da Marambaia e Teresópolis respectivamente apresentam toda a pelagem mais clara, provavelmente devido a um desbotamento provocado pela permanência, durante algum tempo, em formol ou álcool, antes da preparação da pele. O MN 6221 apresenta toda a pelagem mais amarelada, pelos com as bandas amarelas mais pigmentadas (fig. 68).
Comprimento dos pelos: aristiformes, dorsais e laterais 6 mm e ventrais 5 mm; setiformes, dorsais e laterais 4 mm e ventrais 2 mm.
Variação: dos sete exemplares examinados, MN 1308 e MN 1309, da Restinga da Marambaia e Teresópolis respectivamente apresentam toda a pelagem mais clara, provavelmente devido a um desbotamento provocado pela permanência, durante algum tempo, em formol ou álcool, antes da preparação da pele. O MN 6221 apresenta toda a pelagem mais amarelada, pelos com as bandas amarelas mais pigmentadas (fig. 68).
Comprimento dos pelos: aristiformes, dorsais e laterais 6 mm e ventrais 5 mm; setiformes, dorsais e laterais 4 mm e ventrais 2 mm.
Medidas corporais: cabeça e tronco: 77-110 mm; cauda: 43-50 mm; pé: 14-15 mm; orelha: 12-14 mm.

| Monodelphis iheringi, PN Serra dos Órgãos, RJ - foto: Lena Geise |
 |
M. iheringi, foto Thomas Püttker
|
 |
| M. iheringi, foto Thomas Püttker |
Total: 21. BRASIL - Rio de Janeiro: Ilha Grande, 2 M (MZUSP 1979, MN 24400); Jacarepaguá, 1 M (MN 24545); Parati, 2 M (MN 8203, 6221), 2 ? (MN s/n, s/n); Restinga da Marambaia, 1 M (MN 1308); Rio de Janeiro, 1 M (MN 10305), 1 F (MN 24546), 1 ? ( MN s/n); Teresópolis, 1 F (MZUSP 1855). - São Paulo: Iguape ?. - Santa Catarina: Colônia Hansa, 3 M (MZUSP 847, 873, 874); Hammonia, 1 M (MZUSP 3417); Joinville, 2 M (MZUSP 3421(a), 3421(b); 1 ? (MZUSP MB 2).
 |
| Monodelphis gardneri Solari, Pacheco, Vivar and Emmons, 2012 |
Descrição original em inglês:
Holotype.—MUSM 24216, a skin, skull, and alcoholic carcass, all in good condition, of an adult female caught by Elena Vivar (field number EV 1570) on 11 Sep 2007.
Type locality.—Abra Esperanza, San Alberto (Fig. 1, black star), Oxapampa Province, Pasco Department, Per´u, elev. 2784 m; GPS coordinates, taken in the field, are 118560S, 718170W.
Paratypes.—We designate seven specimens as paratypes: one trapped near the type locality, at Estación ‘‘El Cedro’’, San Alberto, 2400 m (MUSM 11334, adult male, fluid specimen); three from the eastern slope of the Cordillera Carpish, Hua´ nuco Department (LSUMZ 14019, adult male, skin, skull and skeleton; MUSM 18943, adult male, fluid specimen; 18944, sub-adult female, skin and skull); two from Cordillera Vilcabamba, 2050 m, Jun´ın Department (MUSM 13007, adult female, skin, skull and alcoholic carcass; USNM 582109, adult male, skin and skull); and one from La Convenci´on, Wayrapata, 2445 m, Cusco Department (MUSM 14631, juvenile female, skin, skull and skeleton).
Distribution.—Montane forests on the eastern slope of the Andes in central and southern Per´u (Fig. 1, black diamonds). At present, M. gardneri is known from the departments of Hua´ nuco, Pasco, Jun´ın, and Cuzco, from 1785 to 2800 m elevation.
Diagnosis.—A species of Monodelphis characterized by three conspicuous blackish stripes on the back; median stripe extending from between ears to base of tail, wider at mid-back, with two shorter, prominent lateral stripes extending from shoulders to base of tail. Size small, especially as reflected in head-and-body length (76–100 mm) and condylobasal length (21.8–27 mm); tail absolutely short (42.5–53 mm) but relatively long (LT 49–52% of HBL). Braincase globular; infraorbital foramen dorsal to M1; alisphenoid portion of bulla relatively inflated, with anterior elongated projection or alisphenoid strut forming a secondary foramen ovale (see Voss & Jansa 2003); (primary) foramen ovale almost entirely limited by alisphenoid, the petrosal delimiting the narrow posterior end. No contact between premolars, either upper or lower; molars small and delicate, with poorly developed ectoflexi on M1 but well-developed on M3–M4.
External measurements in mm of the holotype (and range of three paratypes): HBL 126 (93–149), LT 42.5 (39–52), HF 15.5 (14–15), E 10 (12–14); W 10.25 g.
 |
| Monodelphis gardneri, Pampa Hermosa River, San Ramón, Peru |
 |
| Monodelphis gardneri, Villa Rica-Oxapampa road, Pasco, Peru |
 |
| Monodelphis gardneri, Vilcabamba, Peru |
Description.—Monodelphis gardneri is a small species (HBL , 150 mm), with short dorsal fur (4 mm); Prout Brown to Mummy Brown, grizzled in dorsal view, three conspicuous blackish dorsal stripes (Fig. 3). Median stripe reaching from between ears to base of tail, widest at mid-back; lateral stripes equally evident, but shorter, extending from shoulders to base of tail. Rump tinged rufous in most specimens, but not conspicuously so (rufous expression may be age related). Rhinarium lackish; pinnae sparsely covered by fine short hairs, appearing macroscopically naked, dark brown, blackish in juvenile. Underparts are Brussels Brown in adults, but Mummy Brown in juveniles; no evidence of throat gland. Tail slightly shorter than half of the head-and-body length; but relatively longer than in similar species (see Comparisons); only slightly paler ventrally; well covered by dense and dark scale-hairs. Dorsal surface of manus and pes well furred, but fingers and toes may appear naked at first glance.
Skull small, braincase globular and not flattened (Fig. 4). Inter- and post-orbital constrictions are evident (Fig. 4a). Braincase lacking parietal ridges or sagittal crest. Anterior projection of premaxillae without rostral process. Posterodorsal projection of premaxillae elongated, interposed between maxilla and nasal, reaching the root of P1; infraorbital foramen dorsal to M1 (Fig. 4c). Maxillopalatine fenestrae elongated, from posterior border of P3 to level of M3 protocone, posterolateral palatal foramina small (Fig. 4b). Alisphenoid portion of bulla well inflated, with anterior elongated projection or alisphenoid strut (Fig. 4b) forming a secondary foramen ovale (see Voss & Jansa 2003, Fig. 8c); (primary) foramen ovale almost entirely limited by alisphenoid, the petrosal delimiting the narrow posterior end (Fig. 4b). Basioccipital short, wide, its anterolateral suture with petrosal a raised crest. Crowns of I2, I3, I4, and I5 increasing in width from front to back; anterior cusps present on P1 and P2; no contact between C1, P1, and P2 (Fig. 4c). Small gaps present between lower premolars. Molars small and delicate, poorly developed ectoflexi on M1 but well-developed on M3–M4.
Comparisons.—Monodelphis gardneri is somewhat smaller than sympatric. peruviana in every external and craniodental measurement but larger than M. osgoodi. Compared to similar looking species, it is smaller than M. americana or M. theresa, except in least interorbital breadth (Table 1) but larger than M. iheringi (measurements reported by Gomes 1991); on average, the tail of M. gardneri is relatively longer although shorter in absolute length (Table 1). Monodelphis americana exhibits a longer, wider, and more defined medial stripe that reaches the top of the head (sometimes to the tip of the nose), a condition that is never present in M. gardneri or M. theresa. Because of the mistaken use of the name M. theresa in previous references to M. gardneri, we emphasize cranial comparisons between it, M. theresa, and M. americana. In dorsal view of the skull, the squamosal portion of the zygomatic arch joins the skull almost perpendicularly in M. gardneri, but this union is slightly oblique (anteriorly oriented) in M. Theresa and M. americana. In ventral view, the maxillopalatine fenestrae are elongated and straight in M. gardneri and M. theresa but slightly concave (to the labial side) in M. americana; the postpalatine torus is straight in M. gardneri and M. americana but slightly arched in M. theresa. The tympanic portion of the auditory bulla is much more inflated in M. gardneri, reaching anteriorly to the base of the zygoma, compared with those of M. americana or M. theresa; the alisphenoid tympanic process elongates to a thin process in M. gardneri, but this forms a medial lamina in M. theresa and M. americana. The foramen ovale is almost concealed between the petrosal and the alisphenoid in M. gardneri, but clearly exposed in M. theresa and M. americana; and finally, the paracanine fossa of M. gardneri reaches the level of the posterior edge of C1 but extends only to its anterior edge in M. theresa and M. americana. The most distinctive characteristic of M. iheringi is its flattened which contrasts with the rounded one braincase, of M. gardneri; the former trait represents a unique condition within the genus.
 |
| Monodelphis scalops Thomas 1888 |
Didelphis (Peramys) scalops Thomas (1888a: 158). Thomas (1888b: 359)
Peramys scalops: Cabrera (1919: 43). Miranda-Ribeiro (1936: 412).
Monodelphis theresa Thomas (1921: 441).
Peramys therezae: Miranda-Ribeiro (1936: 416).
Monodelphis iheringi: Vieira (1949: 359) parte.
Monodelphis therezae: Cabrera & Yepes (1940: 31). Vieira (1955: 349).
Monodelphis scalops: Cabrera & Yepes (1940: 32). Davis (1947: 3). Vieira (1949: 360). Vieira (1955: 349) parte. Pine &Abravaya (1978: 379). Emmons (1990: 28).
Monodelphis americana: Davis (1947: 3).
Monodelphis (Monodelphis) americana theresa: Cabrera (1958: 7).
Monodelphis (Monodelphis) scalops: Cabrera (1958: 9).
Monodelphis scalops: Pine & Abravaya (1978: 379).
[Monodelphis] theresa: Pine & Handley, Jr (1984: 242).
Monodelphis scalops: Pine & Handley, Jr (1984: 242).
Monodelphis scalops: Emmons (1990: 28).
Monodelphis theresa: Emmons (1990: 30) parte.
Localidade tipo:" Brasil " restrita para Rio de Janeiro, Teresópolis, por Vieira (1949).
Esta espécie foi descrita por Thomas (1888a: 158) com base em um exemplar macho do Brasil. Em sua descrição comparou-a em tamanho a "D[idelphis]. brevicaudata, Erxl.". " A cabeça, o traseiro e a cauda, ruivos avivados; a parte anterior do dorso, os ombros e a barriga, cinzas-olivas grisalhos. Crânio comprido e estreito; dentes muito pequenos. Medidas: corpo: 133,0; cauda: 71,0; orelha: 8,0; comprimento do crânio: 33,0; comprimento M1-M3: 4,9 (em mm). " Thomas (1888b: 359) redescreveu-a citando um segundo exemplar.
Exemplares de Teresópolis, Rio de Janeiro são citados por Miranda-Ribeiro (1936: 412).
Thomas (1921: 441) descreveu Monodelphis theresa, com base em uma fêmea adulta de Teresópolis, Serra dos Órgãos, Brasil. Considerou-a mais relaciona a M. iheringi. "Tamanho como em M. iheringi, pelagem igualmente curta e fechada, pêlos dorsais com quatro milímetros. Coloração semelhante, porém a cor geral cinza grisalho escuro, nos dois terços anteriores do corpo; a cabeça e o traseiro ruivos-acastanhados vivos. Linhas dorsais quase obsoletas, interrompida na cabeça e mal definida no traseiro, terminando nas ancas; linhas laterais com cerca de uma polegada de comprimento, mal definidas e pouco perceptíveis. Superfície ventral cinza manchado, pêlos com pontas brancas opacas. Bochechas e topo da cabeça ruivo vivo; queixo ruivo pálido. Mãos marrons. Pés externamente marrons e internamente mais esbranquiçados opacos; dedos nus. Cauda marrom no dorso e clareando ventralmente. Crânio não tão achatado como em M. iheringi, mais como em M. americana, caixa craniana comparativamente alta e arredondada. Medidas: corpo: 80,0; cauda: 36; orelha: 10,0; comprimento da crânio: 25,2; largura zigomática: 5,4; comprimento M1-M3: 4,6 (em mm). Distingue-se de M. iheringi por sua cabeça e traseiro ruivos, patas anteriores do dorso acinzentado e linhas dorsais pouco perceptíveis".
Esses dois nomes foram mantidos separados como duas espécies por Miranda-Ribeiro (1936: 416), Cabrera (1940: 31-32), Vieira (1949: 359-360 e 1955: 349), Davis (1947: 3). Cabrera (1958: 7) criou Monodelphis (Monodelphis) americana theresa e manteve Monodelphis (Monodelphis) scalops separado.
Pine & Abravaya (1978: 379) em virtude da aquisição de três exemplares machos de Santa Teresa, Espírito Santo, publicam notas sobre o material novo e comparam-no com as descrições de Thomas, fornecendo um histórico da espécie.
Pine & Handley, Jr. (1984: 242) consideram [Monodelphis] scalops e [Monodelphis] theresa como espécie válidas, ignorando Cabrera. Emmons (1990: 28 e 30) obedece a mesma orientação.
Os dezoito exemplares examinados por mim, permitiram a sinonimização dessas duas espécies tratadas. Principalmente o material do Museu Nacional do Rio de Janeiro, coletado em Teresópolis, RJ. Esses exemplares apresentam idade variada e compreendem cinco machos e quatro fêmeas. O material do Espírito Santo, um macho e três fêmeas também foi decisivo na compreensão do fato.
O padrão da pelagem difere com a idade, os jovens apresentam três faixas dorsais nítidas como em Monodelphis americana, e com o idade essas faixas desaparecem gradativamente. Uma seqüência perfeita pode ser vista nos nove exemplares de Teresópolis.
Na Fazenda Bôa Fé, em Teresópolis, foram coletados três machos, duas fêmeas de M. scalops e uma fêmea de Monodelphis americana. Também em Teresópolis foi coletada M. iheringi, que ocorre em outras localidades junto com M. scalops, como Pedra Branca em Parati e na cidade do Rio de Janeiro, ambas no Estado do Rio de Janeiro.
Davis (1947: 3) cita um curioso incidente notado por um dos coletores. "Logo depois de armar uma ratoeira com banana, ele ouviu esta desarmar e voltou ao local onde encontrou um Monodelphis scalops na ratoeira e um M. americana correndo ao seu redor. Ele removeu o marsupial e armou novamente a ratoeira enquanto o outro permaneceu aproximadamente a um metro de distância. Este imediatamente retornou e foi apanhado."
No material do Museu Nacional, examinei um macho de M. scalops (MN 7245) de Teresópolis (Fazenda Bôa Fé), RJ, coletado no dia 11 de outubro de 1942 por Galdino Pereira 14875 para o Serviço de Pesquisa da Febre Amarela. Examinei também uma fêmea de M. scalops com três faixas dorsais (MN 7249) coletada na mesma localidade e data, pelo mesmo coletor com o número de campo 14876 para o EPFA, ou seja o número seguinte ao do exemplar macho. Os exemplares citados por Davis em sua nota, e não correspondem a espécie diferentes mas sim a M. scalops, talvez um casal.
Ruschi (1948: 3) cita em sua "Lista dos mamíferos do estado do Espírito Santo", a presença de "Monodelphis iheringii [sic] (Thomas)" em Castelo e Alegre, é possível que os animais assim identificados, representem jovens de M. scalops, já coletada em localidades próximas.
 |
| Monodelphis scalops - IPBio, Reserva Betary |
Descrição da pelagem
Coloração geral cinza e marrom avermelhado, partes superiores da cabeça até a nuca, focinho, bochechas, ao redor das orelhas e partes superiores dos membros anteriores, alaranjado, bem vivo no focinho, bochechas e na frente das orelhas; topo da cabeça com pêlos pretos misturados com pêlos ruivos; partes superior e laterais do pescoço, metade anterior do dorso e das laterais, cinza grisalho leve-mente amarelado, pêlos aristiformes dorsais com banda basal e banda terminal preta, pêlos setiformes, dorsais com banda basal cinza, banda subterminal branco amarelado e ponta preta; pêlos aristiformes laterais com banda basal cinza, banda subterminal sépia e banda terminal branco levemente amarelado, pêlos setiformes laterais com banda basal cinza, banda subterminal sépia e ponta branco amarelado; metade posterior do dorso e das laterais, traseiro, partes superiores dos membros posteriores, parte superior da cauda, marrom avermelhado vivo, pêlos setiformes com banda basal cinza, subterminal laranja e ponta preta; cinza e marrom avermelhado dorsais se interpenetrando gradativamente; queixo, bordas da boca e garganta, alaranjado claro, pêlos sem banda basal cinza; parte inferior do pescoço, peito, barriga e partes internas dos membros, cinza levemente amarelado, pêlos com banda basal cinza e banda terminal amarelo claro; parte superior das patas, marrom alaranjado; plantas das patas posteriores, enegrecidas; plantas das patas anteriores claras; cauda com pêlos iguais ao corpo apenas na base, o restante com escamas cobertas por pequenos pêlos, mais longos dorsalmente; parte inferior da cauda, marrom alaranjado claro (fig. 68).
Variação: A coloração da cabeça varia entre marrom alaranjado e ruivo. A metade anterior do dorso varia entre cinza grisalho e cinza grisalho amarelado. O marrom avermelhado da metade posterior do dorso varia entre bem avermelhado e vivo até levemente avermelhado. O avermelhamento é mais intenso no traseiro e na região central do dorso. O exemplar MN 7246 apresenta três estrias longitudinais mais avermelhadas na metade posterior do dorso, no MN 7247 também podem ser notadas muito fracamente. Coloração ventral variando entre cinza levemente amarelado a fortemente amarelado, devido a pêlos com banda terminal estreita ou larga.
Comprimento dos pelos: aristiformes, dorsais 7 mm, laterais e ventrais 8 mm, setiformes, dorsais e laterais 6 mm, ventrais 4 mm.
Medidas corporais: cabeça e tronco: 115-165 mm; cauda: 50-71 mm; pé: 16-25 mm; orelha: 11-16 mm.
Descrição dos jovens e adultos jovens
Coloração geral marrom acinzentado com estrias dorsais. Partes dorsal e laterais da cabeça e membros anteriores, marrom alaranjado mais vivo ao redor das orelhas e bochechas; pescoço dorsal e lateralmente, dois terços anteriores do dorso e laterais, marrom acinzentado, levemente alaranjado lateralmente, pêlos aristiformes dorsais e laterais com banda basal cinza e terminal sépia, nas laterais e com banda basal cinza, subterminal branco amarelado e ponta sépia, pêlos setiformes laterais com banda basal cinza, subterminal amarelo e ponta sépia; parte posterior do dorso e das laterais, partes superiores dos membros posteriores, traseiro e base da cauda, marrom avermelhado escuro; três faixas negras dorsais, a central começando no focinho e diluíndo-se entre os olhos, no topo da cabeça e recomeçando na nuca, as laterais começando nos ombros, todas três evidentes apenas na porção central do dorso e bem diluídas anteriormente e no traseiro; queixo, bordas inferiores da boca e garganta, alaranjado claro; partes inferior do pescoço, peito, barriga e partes internas dos membros, cinza levemente amarelado, pêlos aristiformes com banda basal cinza, mediana marrom e terminal branco acinzentado, pêlos setiformes com banda basal cinza e terminal branco acinzentado; partes superiores das patas, marrom; plantas das patas enegrecidas, patas anteriores mais claras; cauda com pêlos iguais aos do corpo apenas na base, o restante com escamas parcialmente cobertas por pequenos pêlos, marrom escuro em cima, marrom claro em baixo (fig. 68).
 |
| jovens de Monodelphis scalops |
Variação: a coloração dorsal é composta de pelos setiformes de banda basal cinza, banda subterminal amarela e ponta preta. A banda subterminal pode variar entre amarelo bem claro até laranja forte. O exemplar subadulto MN 7249, tem as bandas subterminais amarelo bem claro nos dois terços anteriores do corpo, resultando num cinza levemente amarronzado. O MN 7248 tem bandas subterminais alaranjadas por todo o corpo resultando um marrom alaranjado geral, cabeça, bochechas, ao redor das orelhas, partes superiores dos membros, laterais do corpo e traseiro mais alaranjado que nos outros exemplares. As faixas podem ser pretas, marrom ou cinzentas. No MN 7249 são muito claras, mal destacadas do fundo cinzento. O MN 7233 apresenta laterais mal definidas e acinzentadas e a central evidente de cor sépia. Da mesma forma que no dorso as bandas terminais dos pêlos ventrais variando no comprimento e na coloração, originando desde cinza escuro e cinza claro a cinza alaranjado. O MN 7249 subadulto tem a coloração ventral cinza escuro, a banda basal cinza‚ mais larga que a ponta amarelo claro.
Comprimento dos pelos: aristiformes, dorsais 6 mm, laterais 8 mm e ventrais 7 mm, setiformes, dorsais e laterais 5mm e ventrais 4 mm.
Crânios comparação dos adultos velhos e adultos jovens.
 |
| Dentes selecionados de Monodelphis scalops |
 |
| M. scalops, algumas características anatômicas |
Espécimes examinados
Total: 18. BRASIL - Espírito Santo: Alfredo Chaves, 1 F (MZUSP 6674); Venda Nova, 1 M (UFPB 263), 2 F (UFPB 262, 264); - Rio de Janeiro: Parati, 1 M (MN 6419), 1 F (MN 6102); Petrópolis, 1 F (MN 10209); Rio de Janeiro, 1 F (MN 24546); Teresópolis, 5 M (MN 1301, 1302, 7245, 7246, 7247), 4 F (MN 1310, 7233, 7248, 7249); - São Paulo: São Sebastião, 1 F (MZUSP 1528).

LINKS
Monodelphis scalops
:https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEgaYPYIxybesu9cvAOJ0c6hUQYDDyTXmqL38GqykzCdYwgrGOOZhE50unByYVYGWBqJdIyB9ygfjY9DaJGiF3Ewj2ECEvyim0O0Uq00QTwVxbwvM_xhWHho3W5SyGhGEjvQ7cDwJRm8AqvQ/s1600/MonoIhe2.jpg
Material examinado:
Adultos de Monodelphis scalops

COMPARAÇÃO ETÁRIA :
 | ||
Esse desaparecimento das listas nos adultos lembra o mimetismo dos cervídeos, das antas, dos javalis, dos pumas e leões. Também no caso de Monodelphis pode estar relacionado ao eritrismo que ocorre nos machos velhos de M.americana.
 |

 |
| Monodelphis scalops, Foz do Iguaçu, Paraná - BRASIL foto: Lucas Ferreira |
 | ||
| Monodelphis scalops, Puerto Iguazú, Misiones, ARG Foto: (c) Marcelo Cavicchia
|

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
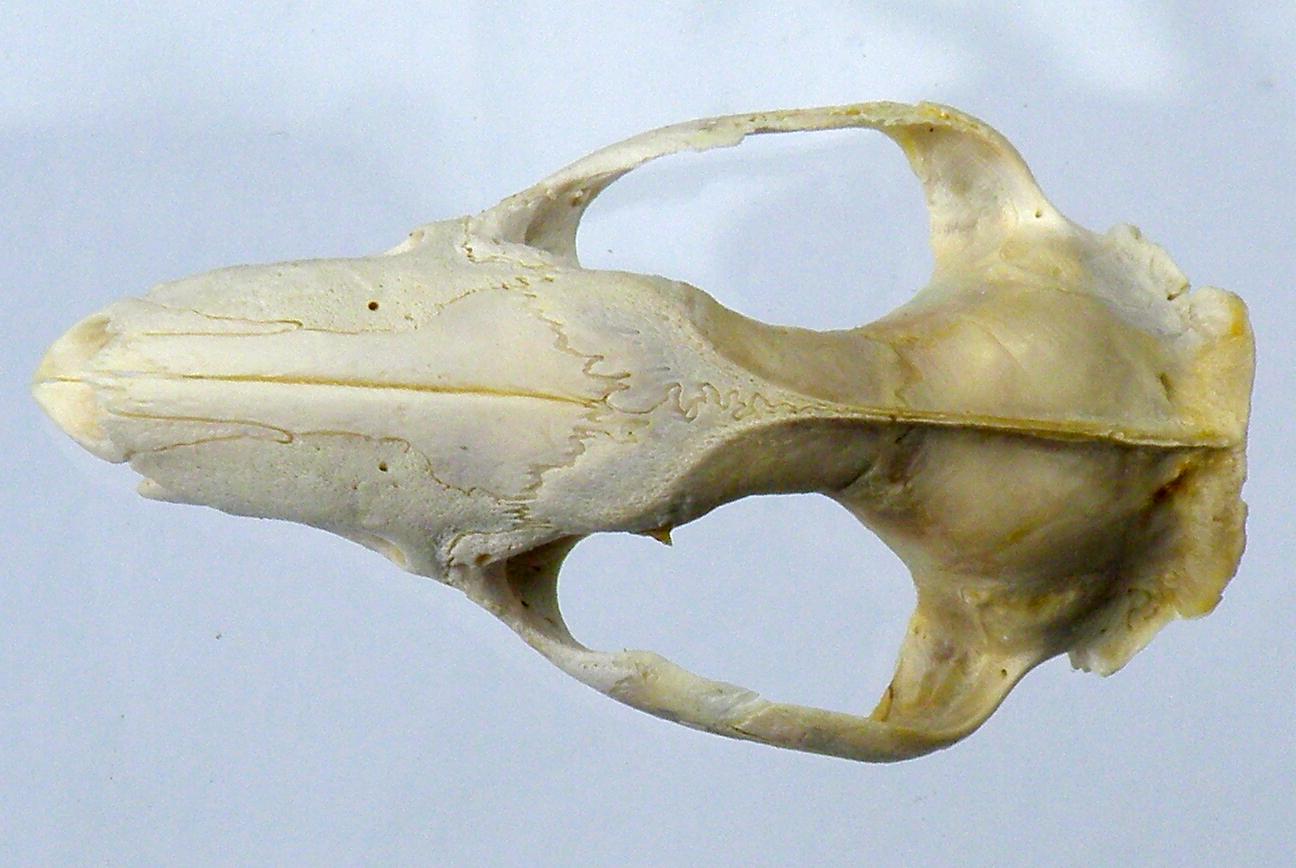{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 |
| Monodelphis scalops, SP, Brasil - foto: Fabiana Umetsu |